C-vitamiini: antioksidantti
Redox-metabolia ja C-vitamiinin antioksidanttiominaisuudet
vapailla radikaaleilla ja hapettimilla on kaksoisrooli sekä myrkyllisinä että hyödyllisinä yhdisteinä aineenvaihdunnassa ja vasteena eksogeenisille stimulaatioille. Ne syntyvät joko normaalista aineenvaihdunnasta tai ympäristötekijöistä (saastuminen, tupakansavu ja säteily). Kun vapaiden radikaalien ylikuormitusta ei voida raadella, niiden kertyminen elimistöön aiheuttaa oksidatiivista stressiä . Oksidatiivinen stressi syntyy, kun vapaiden radikaalien muodostuminen ylittää niitä vastaan suojautumiskyvyn. Tämä prosessi johtaa kroonisten ja rappeuttavien sairauksien, kuten syövän, autoimmuunisairauksien, ikääntymisen, kaihin, nivelreuman, sydän-ja verisuonisairauksien ja hermoston rappeutumissairauksien kehittymiseen . Antioksidantti on molekyyli, joka estää muiden molekyylien hapettumista. Hapetusprosessi on kemiallinen reaktio, joka tuottaa vapaita radikaaleja, mikä johtaa ketjureaktioihin, jotka voivat vaurioittaa soluja. C-vitamiinin antioksidanttivaikutus on hyvin dokumentoitu . C-vitamiini on voimakas antioksidantti, jolla on kyky luovuttaa vetyatomi ja muodostaa suhteellisen vakaa askorbyylivapaa radikaali. E-vitamiini, C-vitamiini ja β-karoteeni tunnetaan antioksidantti vitamiineja, joiden ehdotetaan vähentää oksidatiivisia vaurioita ja alentaa riskiä tiettyjen kroonisten sairauksien. Sairaudet, kuten sydän-ja verisuonisairaudet, liittyvät epidemiologisissa tutkimuksissa riittämättömiin l-askorbiinihapon, tokoferolin ja β-karoteenin pitoisuuksiin. C-vitamiini parantaa myös raudan imeytymistä pelkistämällä Fe3+: n Fe2+: ksi muista kuin hemirautalähteistä . Redox-aktiivisten ionien (rauta, kupari) läsnä ollessa C-vitamiini toimii prooksidanttina ja edistää hydroksyyliradikaalien muodostumista, mikä voi johtaa lipidiin, DNA: han tai proteiinin hapettumiseen . Oksidatiivisen stressin lievittämiseen ja vaurioituneiden makromolekyylien korjaamiseen on olemassa erilaisia mekanismeja. Entsymaattisilla ja ei-bentsymaattisilla antioksidanteilla on tärkeä rooli vapaiden radikaalien ja reaktiivisten happilajien (Ros) keräämisessä. Oksidatiivisen stressin on osoitettu vaikuttavan merkittävästi antioksidanttientsyymeihin katalaasi (CAT), superoksididismutaasi (SOD), glutationireduktaasi (GR), glutationiperoksidaasi (GSHPX) ja kasveissa askorbaattiperoksidaasi (AA-px) ja ei-bentsymaattiset antioksidantit, mukaan lukien glutationi (GSH) ja askorbaatti (ASC). Antioksidanttiyhdisteet voivat estää vapaiden radikaalien hallitsemattoman muodostumisen tai estää niiden reaktion biologisilla sivustoilla; myös useimpien vapaiden radikaalien tuhoutuminen riippuu endogeenisten antioksidanttien hapettumisesta pääasiassa raadannan ja pelkistävien molekyylien avulla . C-vitamiinin ajatellaan olevan tärkeä vesiliukoinen antioksidantti, jonka kerrotaan neutraloivan ROS: ia ja vähentävän oksidatiivista stressiä .
C-vitamiini on voimakas pelkistäjä ja vapaiden radikaalien haaskaaja biologisissa järjestelmissä . Se on mukana antioksidanttipuolustuksen ensimmäisessä linjassa, suojaten lipidikalvoja ja proteiineja oksidatiivisilta vaurioilta. Vesiliukoisena molekyylinä C-vitamiini voi toimia sekä solujen sisällä että ulkopuolella, ja se voi neutraloida vapaita radikaaleja ja estää vapaiden radikaalien vaurioita. C-vitamiini on erinomainen elektronien lähde vapaille radikaaleille, jotka etsivät elektronia saadakseen vakautensa takaisin. C-vitamiini voi luovuttaa elektroneja vapaille radikaaleille ja sammuttaa niiden reaktiivisuuden .
C-vitamiinin on osoitettu olevan tehokas happi-ja typpioksidilajeja, kuten superoksidiradikaali-Ionia, vetyperoksidia, hydroksyyliradikaalia ja singlettihappea vastaan. Tällä C-vitamiinin ominaisuudella on elintärkeitä prosesseja solujen komponenttien suojaamisessa vapaiden radikaalien aiheuttamilta vaurioilta. Lisäksi C-vitamiini uudistaa tehokkaasti E-vitamiinin antioksidanttimuotoa vähentämällä tokoferoksyyliradikaaleja. Tämä prosessi suojaa solukalvoja ja muita lokeroita vapaiden radikaalien aiheuttamilta vaurioilta (kuva 2). Askorbaattiperoksidaasi (APX) on entsyymi, joka pelkistää H2O2: n vedeksi käyttämällä askorbaattia elektronien luovuttajana. Monodehydroaskorbaatti on hapettunut askorbaatti, joka regeneroituu monodehydroaskorbaattireduktaasin (mdar) avulla. Monodehydroaskorbaattiradikaali disproportioituu nopeasti askorbaatiksi ja dehydroaskorbaatiksi. Dehydroascorbate is reduced to ascorbate by dehydroascorbate reductase in the presence of GSH, yielding oxidized glutathione (GSSG). It is reduced by glutathione reductase (GR) using nicotinamide adenine dinucleotide phosphate hydrogen (NADPH) as an electron donor. Dehydroascorbate may be reduced nonenzymatically or catalyzed by proteins with dehydroascorbate reductase (DHAR) activity.
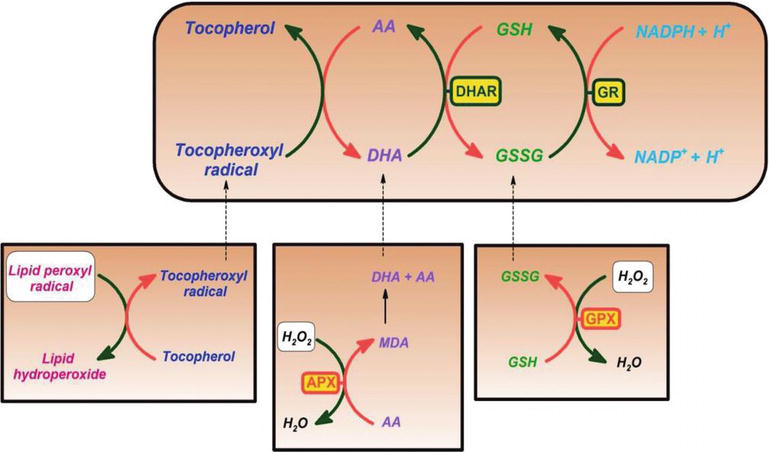
Figure 2.
xmlns:mml=”http://www.w3.org/1998/Math/MathML” xmlns:xlink=”http://www.w3.org/1999/xlink” xmlns:xsi=”http://www.w3.org/2001/XMLSchema-instanceAscorbate and redox cycling antioxidants. AA, ascorbate; DHA, dehydroascorbate; DHAR, semidehydroascorbate reductase; GSH, glutathione; GSSG, semi-glutathione reductase; GR, glutathione reductase; APX, ascorbate peroxidase; and GPX, glutathione peroxidase .
Glutathione-ascorbate cycle operates in the cytosol, mitochondria, plastids, and peroxisomes in plants . On ehdotettu, että glutationi-askorbaattisykli on avainasemassa H2O2: n detoksifikaatiossa, koska kasvisoluissa on suuria glutationin, askorbaatin ja NADPH: n pitoisuuksia. Myös muut entsyymit, kuten askorbaatti ja glutationiperoksidaasit, jotka käyttävät tioredoksiineja tai glutaredoksiineja pelkistävinä substraatteina, osallistuvat H2O2: n poistoon kasveissa (kuva 2).
C-vitamiini muodostaa myös semidehydroaskorbyyliradikaalin, suhteellisen pitkäikäisen radikaalin, regeneroidessaan E-vitamiinia sen radikaalimuodosta, samoin kuin haaskaradikaaleissa. Kasvi-ja eläinsoluissa on NADH-riippuvainen semidehydroaskorbaattireduktaasientsyymi (EC 1.6.5.4), joka pelkistää radikaalin takaisin C-vitamiiniksi käyttämällä pelkistimenä NADH: ta (kuva 2). Sekä entsymaattisesti että ei-bentsymaattisesti se voi hajota peruuttamattomasti diketoglukonihapoksi tai se voidaan muuttaa askorbaatiksi glutationiriippuvaisessa reaktiossa .
pelkistävänä aineena ja elektronien luovuttajana C-vitamiini luovuttaa vapaiden radikaalien keräämisessä korkeaenergisiä elektroneja vapaiden radikaalien neutraloimiseksi, ja se hapettuu dehydroaskorbiinihapoksi. Dehydroaskorbiinihappo voidaan muuttaa takaisin askorbiinihapoksi uudelleenkäyttöä varten tai se voi metaboloitua vapauttaen edelleen Lisää elektroneja. Vaikka C-vitamiini imeytyy suolesta natriumriippuvaisen C-vitamiinin kuljettajaproteiinin kautta, useimmat solut kuljettavat C-vitamiinia hapettuneessa muodossa (dehydroaskorbiinihappo) glukoosin kuljettajaproteiini 1: n kautta. Dehydroaskorbiinihappo pelkistyy tuottamaan askorbiinihappoa solun sisälle, mikä suojaa mitokondrioita vapaiden radikaalien aiheuttamilta oksidatiivisilta vaurioilta (Kuvat 2 ja 3). Erittäin reaktiiviset vapaat radikaalit (esim., RO–, RO2–, OH -,, NO2) pelkistyvät askorbaatilla, ja juuri syntynyt askorbyyliradikaali on huonosti reaktiivinen. Askorbaatti voi myös haaskata peroksinitriitistä johdettuja ei-radikaalisia reaktiivisia lajeja, kuten hypokloorihappoa, otsonia ja nitraavia aineita. C-vitamiini on monosakkaridien hapetus-pelkistyskatalyytti (redox), jota esiintyy sekä eläimillä että kasveilla. C-vitamiinin antioksidanttivaikutus johtuu sen kyvystä luovuttaa elektroneja sekä toisesta että kolmannesta hiilestä. Kädellisten evoluution aikana yksi askorbiinihapon valmistukseen tarvittavista entsyymeistä on kadonnut mutaation seurauksena, ihmisten on saatava se ruokavaliosta; useimmat eläimet voivat syntetisoida tätä vitamiinia kehossaan eivätkä tarvitse sitä ruokavaliossaan . C-vitamiinia tarvitaan prokollageenin muuntamisessa kollageeniksi hapettamalla proliinijäämät hydroksiproliiniksi. Muissa soluissa se säilyy pelkistetyssä muodossaan reaktiolla glutationin kanssa . Kuten kuvioista 2 ja 3 ilmenee, askorbiinihappo on redox-katalyytti, joka voi pelkistää ja siten neutraloida ROS: ää, kuten vetyperoksidia (H2O2) (Kuvat 2 ja 3).
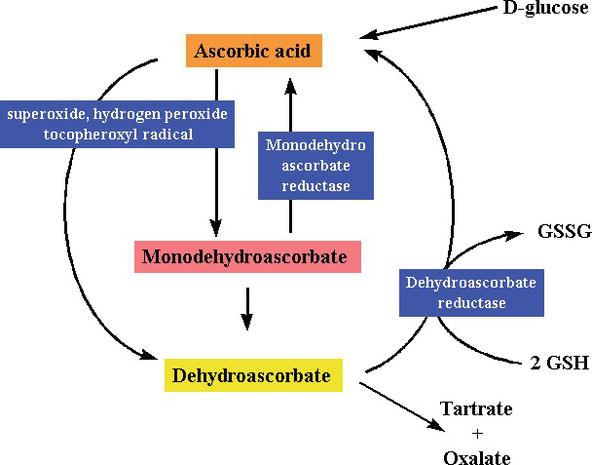
Figure 3.
xmlns:mml=”http://www.w3.org/1998/Math/MathML” xmlns:xlink=”http://www.w3.org/1999/xlink” xmlns:xsi=”http://www.w3.org/2001/XMLSchema-instanceSynthesis and degradation ofl-ascorbic acid in plant tissues .
Ascorbic acid has direct antioxidant effects, and also it is a substrate for the redox enzyme ascorbate peroxidase, that is particularly important in stress resistance in plants. Askorbiinihappoa esiintyy runsaasti kaikissa kasvinosissa, erityisesti kloroplasteissa, jotka saavuttavat siellä 20 millimetrin pitoisuudet . Dehydroaskorbaatti (DHA) ja askorbaatti vapaa radikaali (AFR), välituotteena askorbaatti vapaa radikaali (AFR), jotka ovat palautuvia, yhden elektronin hapetuksia syntyy askorbaatista (Kuva 4). Yleisesti oletetun ROS: n entsymaattisen poistomallin mukaan SOD katalysoi superoksidianionin H2O2: ksi ja hapeksi, minkä jälkeen H2O2 pelkistyy vedeksi ja molekyylihapeksi CAT: n avulla. CAT turnover number is very high, but its affinity for H2O2 is relatively low, and consequently a certain amount of H2O2 remains in the cell.
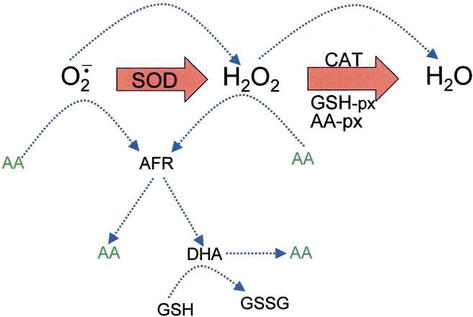
Figure 4.
xmlns:mml=”http://www.w3.org/1998/Math/MathML” xmlns:xlink=”http://www.w3.org/1999/xlink” xmlns:xsi=”http://www.w3.org/2001/XMLSchema-instanceThe role of AA in the detoxification of ROS. Blue dotted lines indicate nonenzymatic reactions.
H2O2 can react with superoxide anion formed in oxidative metabolism generating the highly reactive hydroxyl radical. GSH peroxidases (GSH-px) and AA peroxidases (AA-px) are capable of scavenging H2O2 due to their high affinity for H2O2. The cooperativity of SOD, CAT, and peroxidases ensures low amounts of superoxide anion and H2O2 and limiting the risk of hydroxyl radical formation (Figure 5).

Figure 5.
xmlns:mml=”http://www.w3.org/1998/Math/MathML” xmlns:xlink=”http://www.w3.org/1999/xlink” xmlns:xsi=”http://www.w3.org/2001/XMLSchema-instanceFoyer-Halliwell-Asada cycle .