Erityinen sokeri: miten sialiinihappo vaikuttaa aineenvaihduntaan, terveyteen ja sairauksiin
olemme täynnä mikrobeja, jotka elävät kehomme ulko-ja sisäpuolella olevilla pinnoilla. Ymmärryksemme erityisesti suoliston mikrobiomista paranee nopeasti, kun alamme selvittää, miten monimutkaiset mikrobiyhteisöt ovat vuorovaikutuksessa ruokavaliomme, muiden mikrobien ja solupintojemme kanssa. Vaikka monet kemikaalit ovat tärkeitä tässä miljöössä, tässä keskitymme siaalihappoihin kriittisenä molekyylijoukkona, joka tukee monia näistä vuorovaikutuksista ja vaikuttaa aineenvaihduntaan, terveyteen ja sairauksiin.
yleisin sialiinihappo, N-asetyylineuramiinihappo (Neu5Ac) (viikuna. 1) on suhteellisen yksinkertainen sokerihappo, jossa ei ole varaustaan lukuun ottamatta mitään sinänsä epätavallista. Kun solut kuitenkin ottavat yhteyttä muihin soluihin, ne käyttävät usein pinnalla näkyviä molekyylejä, ja tässä siaalihapoista tulee niin tärkeitä, koska ne ovat yleensä isäntäglykaanien terminaalisia sokereita, jotka pippuroivat solupinnoilla istuvia proteiineja ja lipidejä. Solujen pinnalla tai erittämät muciiniglykoproteiinit ovat hyviä esimerkkejä sialiinihappopäällysteisistä rakenteista, joita mikrobit voivat kohdata limakalvopinnoilla, kuten hengitysteiden tai ruoansulatuskanavan alueella. Tämä ainutlaatuinen asema muiden ihmissolujen, bakteerisolujen ja virusten ”meet and greet” -molekyylinä asettaa siaalihapot keskeiselle sijalle monissa tärkeissä prosesseissa.
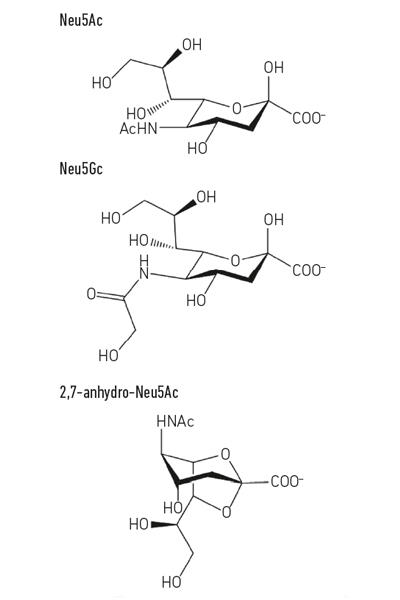
- Kuva. 1. Esimerkki sialihappojohdannaisista. Huomaa, että ihminen ei voi valmistaa n-glykolyyli-neurnamiinihappoa (Neu5Gc), mikä on biokemiallinen ero, joka erottaa meidät muista suurista apinoista, mutta se voidaan sisällyttää solupintoihimme, kun se saadaan ravinnosta.
- sokerikuorrutetut patogeenit
- Ristisyöttö-makeisten jakaminen
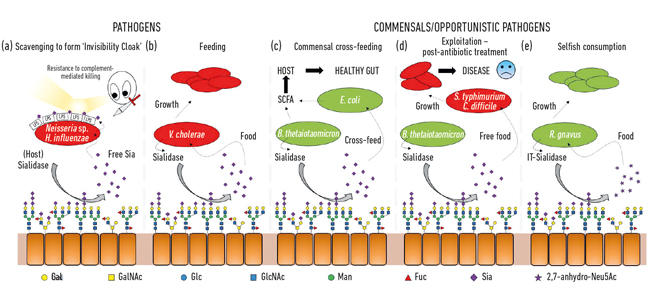
- Kuva. 2. Sarjakuva havainnekuva erilaisista vuorovaikutuksista bakteerien ja sialihappojen läsnä terminaalinen sokerit eri isäntä glykaaneja. Nämä pilkkoutuvat sialidaaseilla ja muuttuvat yleensä vapaiksi Neu5aceiksi lukuun ottamatta IT-sialidaasia, joka vapauttaa 2,7-anhydro-Neu5acia. Taudinaiheuttajat voivat käyttää isäntäeläinten tai patogeenisten sialidaasien vapauttamaa vapaata sialihappoa näkymättömyysviittana (a) tai ruokinnassa (b). Ristisyöttö voi tapahtua suolistomikrobiston jäsenten välillä tai saman lokeron (c, d) jakavien vastaavien ja patogeenisten bakteerien välillä, kun taas muut bakteerit ovat kehittäneet itsekkäitä mekanismeja sialiinihapon varaamiseksi omaan käyttöön (e).
- omista makeisista kiinni pitäminen
- virukset pääsevät mukaan toimintaan
- Jatkoluku
- Andrew Bell
- Emmanuele Severi
- Nathalie Juge
- Gavin H. Thomas
- miksi mikrobiologialla on merkitystä?
- mitä neuvoja antaisit alalla aloittavalle?
Kuva. 1. Esimerkki sialihappojohdannaisista. Huomaa, että ihminen ei voi valmistaa n-glykolyyli-neurnamiinihappoa (Neu5Gc), mikä on biokemiallinen ero, joka erottaa meidät muista suurista apinoista, mutta se voidaan sisällyttää solupintoihimme, kun se saadaan ravinnosta.
sokerikuorrutetut patogeenit
yksi ensimmäisistä sialihappojen mikrobitoiminnoista liittyi joidenkin patogeenien kykyyn kolonisoida ja sitten aiheuttaa sairauksia ihmisillä. Mikrobiologian seuran entinen puheenjohtaja, professori Harry Smith FRS, joka työskentelee Birminghamin yliopistossa, teki uraauurtavaa työtä Neisseria gonorrhoean parissa. Kun hänen ryhmänsä tutkijat eristivät virheen suoraan kehosta, se oli vastustuskykyinen ihmisen seerumin tappamiselle (komplementtivälitteinen tappaminen), mutta jos vikoja viljeltiin laboratoriossa muutaman sukupolven ajan, tämä suoja menetettiin ja solut tapettiin seerumilla. Hänen ryhmänsä oli havainnut, että nämä taudinaiheuttajat varastavat sialiinihappoa isännältä ja päällystävät sillä Oman solupintansa inhimillistääkseen ne. Koska ne voivat kuitenkin vain haaskata tätä sialihappoa, sen puuttuminen kasvualustasta tarkoittaa, että sialiinihappoa ei enää lisätä, joten ne menettävät nopeasti tämän ”seerumisuojan” ja kuolevat. On käynyt ilmi, että joukko muita ovelia vikoja käyttää samanlaisia strategioita oman näkymättömyysviittansa luomiseksi sialiinihapon avulla, kuten Neisseria meningitidis, Campylobacter jejuni tai Haemophilus influenzae (Kuva. 2a)
Ristisyöttö-makeisten jakaminen
sekä taudinaiheuttajien näkymättömyysviittana käyttämä siaalihapot ovat melko hyvää ravintoa bakteereille, sillä niistä saadaan sekä hiiltä että typpeä, ja niitä voidaan käyttää energianlähteenä. Aikaisempi työ osoitti, että Vibrio choleraen kaltaiset taudinaiheuttajat söivät mielellään vain sialihappoja ja että se itsessään oli tärkeää isäntäkolonisaation onnistumiselle (Kuva. 2b).
suolistossa merkittävä sialihappojen lähde tulee mukeista, jotka ovat epiteelin pintaa peittävän limakerroksen päärakennekomponentteja. Kaikkein paljain limakerros on erilaisten yhteismitallisten bakteerien elinympäristö, jotka ovat oppineet herkuttelemaan muciiniproteiineilla, joiden massasta sokerit muodostavat 80 prosenttia. Yksinkertaisesta topologisesta näkökulmasta päätesokereina olevat siaalihapot kerätään ensin talteen, jolloin niiden taustalla olevat sokerit, jotka voidaan myös myöhemmin vapauttaa ja syödä, paljastuvat. Seuraavan sukupolven sekvensoinnin myötä olemme saaneet tietoa tässä lajityypissä elävistä mikrobeista, ja monilla on geenejä sialiinihapon ottoon ja myöhempään kataboliaan. Mielenkiintoista oli se havainto, että jotkin commensal anaerobit, kuten Bacteroides thetaiotaomikron, erittävät sialidaaseja vapauttaakseen sialiinihappoa, mutta niillä ei ole kuljettajia eikä katabolisia geenejä, joiden avulla sitä voitaisiin käyttää. Tämä näennäisen pyyteetön ele tarvitaan, jotta bakteerit pääsisivät käsiksi taustalla oleviin sokereihin ja tukisivat metabolisen ristisyötön prosessia muihin liman lokerossa eläviin ötököihin, kuten vastaaviin Escherichia coli-kantoihin, joilla ei ole sialidaasia, mutta jotka ovat hyviä syömään vapaata sialihappoa (Kuva. 2c). Tämä käsite tunnustetaan nyt tärkeäksi osaksi kolonisaatiota ja lajityyppiin sopeutumista. Nerokkaan esimerkin tästä kuvasi Justin Sonnenburgin ryhmä Stanfordin yliopistosta käyttäen hiirimallia, jossa on yksinkertainen määritelty suolistomikrobisto. Antibioottikuurin jälkeen monet sialihappoa syövistä bakteereista hävisivät, minkä seurauksena vapaata sialihappoa kertyi suolistoon. Kun taudinaiheuttajia, kuten Salmonella typhimurium tai Clostridium difficile, vastaanotetaan, nämä taudinaiheuttajat hyötyisivät vapaa sialiinihappo ravintoaineena (Kuva. 2d). Taudinaiheuttajien mutanttikannoilla, joilta puuttuvat transportterit tai kataboliset geenit, ei ollut etulyöntiasemaa, joten niillä oli suora yhteys taudinaiheuttajan uloskasvuun, kun suoliston mikrobiyhteisö häiriintyy. Siksi ehdotettiinkin mekanismeja vapaan sialiinihapon poolin pienentämiseksi suolistossa osana uutta tartunnanvastaista strategiaa.
Kuva. 2. Sarjakuva havainnekuva erilaisista vuorovaikutuksista bakteerien ja sialihappojen läsnä terminaalinen sokerit eri isäntä glykaaneja. Nämä pilkkoutuvat sialidaaseilla ja muuttuvat yleensä vapaiksi Neu5aceiksi lukuun ottamatta IT-sialidaasia, joka vapauttaa 2,7-anhydro-Neu5acia. Taudinaiheuttajat voivat käyttää isäntäeläinten tai patogeenisten sialidaasien vapauttamaa vapaata sialihappoa näkymättömyysviittana (a) tai ruokinnassa (b). Ristisyöttö voi tapahtua suolistomikrobiston jäsenten välillä tai saman lokeron (c, d) jakavien vastaavien ja patogeenisten bakteerien välillä, kun taas muut bakteerit ovat kehittäneet itsekkäitä mekanismeja sialiinihapon varaamiseksi omaan käyttöön (e).
omista makeisista kiinni pitäminen
vaikka paljolti varhainen sialomikrobiologia keskittyi taudinaiheuttajiin, uudemmassa työssä on keskitytty samassa lajityypissä asuviin vastaaviin bakteereihin. Suolistomikrobiston tämänpäiväiset tutkimukset paljastavat, kuinka kovaa kilpailu neu5acista on suolistossamme ja miten se voi kääntää tasapainon taudinaiheuttajien ja yhteismitallisten välillä. Mitä jos tietty bakteeri voisi vapauttaa sialihappoa muodossa, johon vain se voisi päästä? Työ Juge-ryhmässä Norwichissa osoitti, että suoliston symbiontti Ruminococcus gnavus on kehittynyt tähän, käyttäen erityistä sialidaasia, jota kutsutaan intramolekulaariseksi trans-sialidaasiksi (IT-sialidaasi), joka vapauttaa 2,7-anhydro-Neu5acia (KS.Kuva. 1) sijasta Neu5Ac. Kehittämällä menetelmän puhtaan 2,7-anhydro-Neu5acin syntetisoimiseksi he osoittivat, että R. gnavus-kannat pystyivät kasvamaan 2,7-anhydro-Neu5acilla ainoana hiililähteenä. Entsymaattista reaktiota katalysoivan domeenin lisäksi IT-sialidaasit muodostavat hiilihydraattia sitovan domeenin, joka auttaa välittämään IT-sialidaasin sitoutumista sialihapporikkaisiin muciineihin. In vivo tämä saattaa suosia mekanismia, joka auttaa kohdentamaan bakteerit SIALIHAPPORIKKAISIIN ruoansulatuskanavan alueisiin, mikä edistää bakteerien kolonisaatiota limakerroksessa. Kun se on sijoitettu oikeaan paikkaan, it-sialidaasi pilkkoo sialihapon mukkien terminaaliketjuista ja vapauttaa sen muodossa, 2,7-anhydro-Neu5Ac, että R. gnavus voi käyttää omana ravinnonlähteenään sen sijaan, että jakaisi neu5acin muiden limarakossa asuvien bakteerien, kuten taudinaiheuttajien, kanssa (viikuna. 2 e). Tämä mekanismi tarjoaa kilpailuetua muihin sialihappoja syöviin bakteereihin verrattuna, jotka elävät limarakossa, tarjoamalla ravintoaineen lähteen, jota se voi mieluiten käyttää. Lisäksi R. gnavus voi myös auttaa vähentämään suolistopatogeenien kasvua vähentämällä neu5ac-pitoisuutta limakalvoympäristössä, mikä avaa mahdollisuuksia uudenlaiselle terapeuttiselle strategialle lisääntyvän antibioottiresistenssin aikana.
virukset pääsevät mukaan toimintaan
virukset käyttävät infektiosyklinsä aikana myös sialihappoja avainmolekyyleinä, tunnetuimpana influenssavirus. Alkuperäinen kiinnittyminen on ensimmäinen keskeinen vaihe, kun sialihapot toimivat viruksen reseptoreina hengitysteiden epiteelisoluissa. Sialiinihapon ja glykaanien taustalla olevien sokerien välinen sidos säätelee usein influenssavirusten isäntäaluetta, ja pandemiakannoissa on usein adheesioproteiinia (hemagglutiniinia), joka tunnistaa sialiinihapon kiinnittymisen ihmisen kaltaisiin muotoihin ja on siten erityisen virulentti. Sialiinihappo on tärkeä viruksen elinkaaren viimeisessä vaiheessa, sillä tartunnan saaneesta solusta vapautuminen edellyttää sialihappojen pilkkomista pois isäntäsolun pinnalta. Tätä katalysoi virusentsyymi nimeltä sialidaasi (tai neuraminidaasi). Tamiflu®: ksi (Oseltamiviirifosfaatti) ja Tsanamiviiriksi Muotoillun sialiinihapon rakenteellisten analogien aiheuttama tämän entsyymin esto on ensisijainen hoitomme influenssan hoidossa ja rakenne-/toimintoperusteisen lääkesuunnittelun riemuvoitto.
johtopäätöksenä on selvää, että sialihapot ovat kriittisiä molekyylejä monissa prosesseissa, joissa mikrobisolut ja virukset vuorovaikuttavat nisäkkäiden solupintojen kanssa. Tässä lyhyessä artikkelissa olemme käsitelleet joitakin niistä antaaksemme käsityksen näiden sokerien keskeisistä toiminnoista viestinnässä, häive -, kolonisaatio-ja aineenvaihdunnassa nummen ja sairauksien eri osa-alueilla. Sialihappoja on kuvailtu maailman tärkeimmiksi kemikaaleiksi, ja ne tulevat epäilemättä jatkossakin paljastamaan lisää ihmisten evoluutiosta ja vuorovaikutuksesta mikrobiensa kanssa.
Jatkoluku
Thomas, GH. Sialiinihappo hankinta bakteereissa – yksi substraatti, monia kuljettajia. Biochem Soc Trans 2016;44: 760-765.
ng KM, Ferreyra JA, Higginbottom SK, Lynch JB, Kashyap PC et al. Mikrobien vapauttamat isäntäsokerit helpottavat enteropatogeenien laajenemista antibioottikuurin jälkeen. Nature 2013; 502: 96-99.
Owen CD, Tailford LE, Monaco S, Šuligoj T, Vaux L et al. Selvitetään suoliston Symbiont Ruminococcus gnavus-bakteerin sialiinihapon tunnistamisen spesifisyys ja mekanismi. Nat Commun 2017; 8: 2196.
Tailford LE, Owen CD, Walshaw J, Crost EH, Hardy-Goddard J, Le Gall G, de Vos WM, Taylor GL, Juge N. Discovery of intramolecular trans-sialidases in human gut microbiota viittaa uudenlaisiin limakalvojen sopeutumismekanismeihin. Nat Commun 2015; 6: 7624.
Juge N, Tailford L, Owen CD. Sialidases from gut bacteries: a mini-review. Biochem Soc Trans 2016;44: 166-175.

Andrew Bell
Quadram Institute Bioscience, Norwich Research Park, NR4 7UQ, Iso-Britannia
Andrew Bell on jatko-opiskelija Quadram Instituten Gut Microbes and Health-laitoksella, joka tutkii sialihapon aineenvaihdunnan uuden mekanismin vaikutusta suoliston homeostaasiin. Hän suoritti biokemian BSc-tutkinnon East Anglian yliopistossa ja työskenteli 2 vuotta John Innes Centressä herneen klorofyllimetabolian (Pisum sativum) parissa ennen liittymistään Nathalie Jugen ryhmään Quadram-instituutissa vuonna 2015 tohtorintutkintoa varten.

Emmanuele Severi
Biologian laitos, Yorkin yliopisto, Wentworth Way, York YO10 5DD, Iso-Britannia
@emm_severi
Emmanuele Severi on Prof Gavin Thomasin laboratoriossa työskentelevä post doc-tutkijatohtori Yorkin yliopiston biologian laitoksella. Hän on työskennellyt sialiinihapon kuljetuksen ja aineenvaihdunnan parissa eri aikoina urallaan, ja hän on ollut seuran jäsen vuodesta 2002.

Nathalie Juge
Quadram Institute Bioscience, Norwich Research Park, NR4 7UQ, UK
Nathalie Juge on Norwichissa sijaitsevan Quadram Institute Biosciencen (QIB) tutkimusjohtaja, Institute Strategic Programme, The Gut Microbes and Health (GMH) – ohjelman varajohtaja ja idän yliopiston biologisten tieteiden korkeakoulun kunniaprofessori Anglia. Hän johtaa suoliston isäntämikrobien interaktioiden glykobiologiaa tutkivaa tutkimusryhmää, jonka pääaiheena on mucin-johdannaisen sialiinihapon aineenvaihdunta suoliston mikrobistossa (https://quadram.ac.uk/nathalie-juge).

Gavin H. Thomas
Biologian laitos, Yorkin yliopisto, Wentworth Way, York YO10 5DD, Iso-Britannia
@GavinHThomas
Gavin H. Thomas on mikrobiologian professori Yorkin yliopiston biologian laitoksella ja työskennellyt bakteerien sialiinihapon kuljetuksen ja katabolian parissa yli vuosikymmen. Hän on ollut seuran jäsen vuodesta 1995, oli”Microbiology Today” – lehden päätoimittaja (2003-2006), toimi samalla kaudella valtuustossa, toimi viestintävaliokunnassa (2012-2015) ja tasa-arvovaliokunnassa & Monimuotoisuusvaliokunnassa (2013-2015). Hän on ollut päätoimittaja, vastaava päätoimittaja ja on nykyään seuran johtavan mikrobiologia-lehden (http://thomaslabyork.weebly.com) varapäätoimittaja.
miksi mikrobiologialla on merkitystä?
Gavin: Sillä on merkitystä terveydellemme – vasta nyt alamme todella ymmärtää mikrobiomimme molekyylikoostumusta ja toimintaa, joka todennäköisesti vaikuttaa kehoomme moninaisilla tavoilla.
mitä neuvoja antaisit alalla aloittavalle?
Gavin: Lue laajasti ja pidemmälle ”oma vika” – mikrobit voivat tehdä aika paljon kaikkea ja ovat yleensä keksineet useita eri tapoja tehdä se!
kuvat: H3N2-influenssaviruspartikkelit. CDC / Science Photo Library.
kuva. 1. Andrew Bell.
kuva. 2. Emmanuele Severi.