Helga Kolbin verkkokalvon gliasolut
Helga Kolb
kolmea gliasolun perustyyppiä on ihmisen verkkokalvossa, Mullerin soluissa, astrogliassa ja mikrogliassa. Cajal kuvasi kaikki verkkokalvolle yli sata vuotta sitten (1892).
1. Mullerin sellejä.
Muller-solut ovat verkkokalvon pääasiallinen gliasolu. Ne muodostavat arkkitehtonisia tukirakenteita, jotka ulottuvat säteittäisesti verkkokalvon paksuuden yli ja ovat verkkokalvon rajoja ulko-ja sisärajakalvon kohdalla vastaavasti. Täydellinen ymmärrys Mullerin solun muodosta on parhaiten nähtävissä Golgi-värjäyksen jälkeen, kuten Cajal (1892) alun perin osoitti alla.
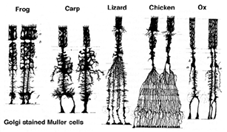
Fig. 1. Golgin värjäämät Mullerin solut
Mullerin solurungot istuvat sisemmässä ydinkerroksessa ja projisoivat epäsäännöllisen paksuja ja ohuita prosesseja kumpaankin suuntaan ulkorajoittavalle kalvolle ja sisemmälle rajoittavalle kalvolle. Mullerin soluprosessit sijoittuvat ydinkerroksissa olevien neuronien solurunkojen ja pleksikerroksissa olevien hermoprosessien kirjekuoriryhmien väliin (Kuva. 1). Itse asiassa verkkokalvon hermoprosessit ovat sallittuja vain suoraan kosketukseen, ilman kuoria Mullerin solujen prosesseja, niiden synapseissa.
yksi kantasolu synnyttää sekä Mullerin soluja että verkkokalvon neuroneja(Turner ja Cepko, 1987), vaikkakin ilmeisesti kahdessa vaiheessa. Varhaisimmat hermosolut, jotka syntyvät pigmenttiepiteelin viereisen neuroepiteelin apikaalisella marginaalilla, tuottavat primaarisia neuroneja, jotka koostuvat kartiosoluista, vaakasuorista soluista ja ganglionisoluista (Fig. 2, oik.). Myös apikaalien reunoilla syntyvien solujen toinen vaihe tuottaa Muller-soluja ja rod-fotoreseptoreita, bipolaarisoluja ja amakriinisoluja (Reichenbach and Robinson, 1995) (Kuva. 2, Vas.). Kaikkien kehittyvien neuronien ja Muller-solujen on siirryttävä sisäänpäin lopulliseen asemaansa, ja on ajateltu, että Muller-solujen prosessit ja rungot ohjaavat suurta osaa hermosolujen vaelluksista ja ohjaavat neuriittien erilaistumista.
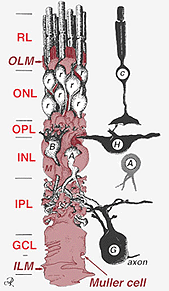
Fig. 2. 3-D-kaavamainen piirustus Muller-solun ja muiden verkkokalvon neuronien suhteesta
ulomman rajoittavan kalvon muodostavat liitokset ovat Muller-solujen ja muiden Muller-solujen ja fotoreseptorisolujen välillä tukevina desmosomeina tai zonula-adhereeneina. Joissakin lajeissa kuilun liitokset (erikoistuneet kalvoliitokset ja kanavat, jotka mahdollistavat pienten molekyylien ja ionien kulun) tai tiiviit liitokset ovat osa näitä Mullerin soluliitoksia (Miller ja Dowling, 1970), mutta näin ei ole nisäkäslajeissa, joissa ei ole koskaan havaittu väriaineliitosta (Robinson et al., 1993; Reichenbach ja Robinson, 1995). Pigmenttiepiteelin ja subretinaalisen tilan edessä olevan Muller-solun pintaa laajentavat monet apikaalisena villinä tunnetun Muller-solukalvon ulokkeet. Sisempi rajauskalvo taas muodostuu Muller-solun kartiomaisesta endfeetistä, mutta tässä ei nähdä erikoistuneita liitoksia. Mullerin solut muodostavat endfeetiä myös verkkokalvon sisäpinnalla oleviin suuriin verkkokalvon verisuoniin. Lasiaiseen päin olevan Mullerin solukalvon pinta on mukopolysakkaridimateriaalin peitossa ja muodostaa siten todellisen kellarikalvon.
Mullerin solut sisältävät glykogeeniä, mitokondrioita ja välivaiheen filamentteja, jotka ovat immunoreatiivisia vimentiiniä ja jossain määrin glial fibrillaarista happoproteiinia (GFAP) vastaan. Jälkimmäiset rihmastot ovat tavallisesti verkkokalvon Muller-solujen sisäosassa ja niiden endfeetissä (viikuna. 3, Vasen), mutta verkkokalvon trauman, kuten verkkokalvon irtoamisen, jälkeen sekä vimentin että GFAP ovat massiivisesti säädeltyjä ja löytyvät koko solusta (Kuva. 3, oikealla) (Guerin et al., 1990; Fisher ja Lewis, 1995).
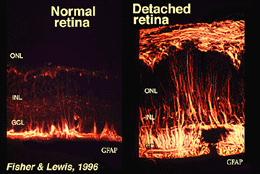
Fig. 3. GFAP-immunoreaktiivisuus Muller-soluissa
Muller-soluilla on joukko toimintoja, jotka kaikki ovat elintärkeitä verkkokalvon neuronien terveydelle. Mullerin solut toimivat symbioottisessa suhteessa neuroneihin (erinomaisesta tarkastelusta katso Reichenbach and Robinson, 1995). Näin ollen Mullerin solujen toimintoja ovat:
- toimittaa anaerobisen aineenvaihdunnan lopputuotteita (glykogeenin hajoamista) aerobisen aineenvaihdunnan polttoaineeksi hermosoluissa.
- he moppaavat hermojätteitä, kuten hiilidioksidia ja ammoniakkia, ja kierrättävät käytettyjä aminohappolähettimiä.
- ne suojaavat hermosoluja altistumiselta ylimääräisille välittäjäaineille, kuten glutamaatille, käyttäen hyvin kehittyneitä kertymämekanismeja tämän lähettimen kierrättämiseksi. Niille on ominaista erityisesti se, että niissä on suuria pitoisuuksia glutamiinisyntaasia.
- ne voivat olla osallisina sekä hermoromun fagosytoosissa että neuroaktiivisten aineiden kuten GABA: n, tauriinin ja dopamiinin vapautumisessa.
- niiden ajatellaan syntetisoivan retinoiinihappoa retinolista (retinoiinihapon tiedetään olevan tärkeä silmän ja hermoston kehityksessä) (Edwards, 1994)
- ne säätelevät homeostaasia ja suojaavat neuroneja ioniympäristön haitallisilta muutoksilta ottamalla solunulkoista K+: aa ja jakamalla sitä uudelleen.
- ne edistävät elektroretinogrammin (Erg) B-aallon (Miller ja Dowling, 1970; Newman ja Odette, 1984), ERG: n hitaan P3-komponentin (Karwoski ja Proenza, 1977) ja scotopic threspondent response (STR) (Frishman and Steinberg, 1989) syntyä. Ne tekevät sen säätelemällä k+ jakautumista verkkokalvon lasiaisen rajan yli, koko verkkokalvolle ja paikallisesti verkkokalvon sisempään pleksikerrokseen (Kuva. 4, Reichenbach ja Robinson, 1995, mukautettu Newman, 1989).

Fig. 4. Mullerin solujen K+: n säätely Mullerin soluissa
2. Astrosyyttejä.
astrosyytit eivät ole verkkokalvon neuroepiteelin gliasoluja, vaan tulevat kehittyvään verkkokalvoon aivoista kehittyvää näköhermoa pitkin (Stone and Dreher, 1987; Chan-Ling, 1994). Niillä on litistyneen solurungon tyypillinen morfologia ja kuitumainen säteilyprosessien sarja. Välivaiheen filamentit täyttävät prosessinsa ja siten ne värjäytyvät dramaattisesti vasta-aineilla GFAP: ia vastaan (Schnitzer, 1988). Astrosyytin solurungot ja prosessit rajoittuvat lähes kokonaan verkkokalvon hermokuitukerrokseen. Niiden morfologia muuttuu periferiasta näköhermon päähän: perifeerisessä verkkokalvossa olevasta symmetrisestä stellaattimuodosta (Figs. 5a ja b) (Schitzer, 1988) erittäin pitkänomaiseen lähelle näköhermoa (Kuva. 6 ja 7).
 Fig. 5A. astrosyytit perifeerisessä verkkokalvossa. Schnitzer, 1988 |
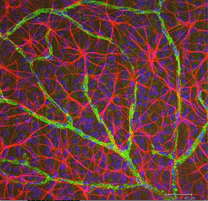 Fig. 5 b. Astrosyytit (punaiset) rotan verkkokalvossa hermosoluissa (siniset) ja verisuonissa (vihreät). Laura Fernandez-Sanchez ja Nicolas Cuenca |
immunosytokemiallisessa värjäyksessä (Kuva. 5B) ja HRP-solunsisäisissä injektioissa (Kuva. 7) värjätyt astrosyytit osoittavat selvästi prosesseja, jotka ovat linjassa hermokuitukerroksen läpi kulkevien hermosolun aksonien kanssa. Jakautumisessa astrosyytit saavuttavat huippunsa näköhermon päässä ja niiden tiheys vähenee melko tasaisesti hermopäästä lähtevissä säteilyrenkaissa. Niitä ei esiinny avaskulaarisessa fovea-tai ora serrata-suonessa.
 Fig. 6. Astrosyyttejä verkkokalvolla. Schnitzer, 1988 |
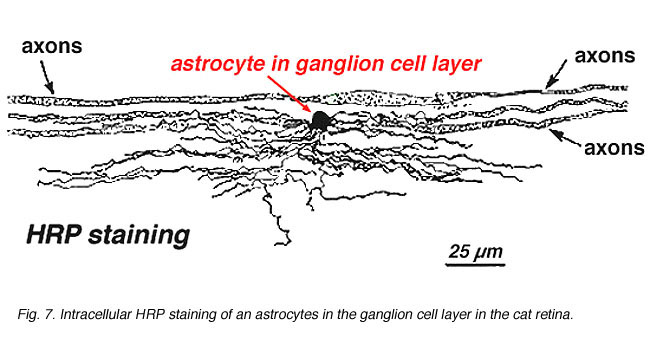 Fig. 7. Astrosyytit kissan verkkokalvossa, jotka on värjätty solunsisäisellä HRP-injektiolla. Kiitos Ralph Nelsonin. |
paksuja ja ohuita astrosyyttejä on erotettu GFAP: n vasta-aineilla tehtyjen värjäysten perusteella (Trevino et al., 1996). Näin astrosyytit ovat järjestäytyneet ganglionsolun aksonikimppujen pinnalle niiden kulkiessa näköhermon päähän muodostaen putken, jonka läpi aksonit kulkevat (Kuva. 8). Gap liitokset ja zonula adherens liitokset on kuvattu astrosyyttisten prosessien välillä cat retina (Höllander et al., 1991).
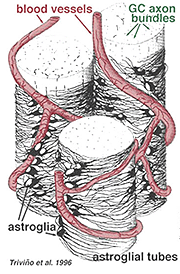
Fig. 8. Astrosyyttien 3-D-lohko, joka on järjestetty ganglion-solun aksonikimppujen pinnalle
, kattaa myös molemmat prosessit ja jopa astrosyytin satunnaisen solurungon. Ganglionisoluaksoneja ympäröivien astrosyyttien toiminta ja suhde hermokuitukerroksen verisuoniin viittaa siihen, että ne ovat aksonaalisia ja vaskulaarisia gliasuojia ja osa veri-aivoestettä. Samoin kuin Muller-solut, niiden tiedetään sisältävän runsaasti glykogeenia ja ne voivat muodostaa ravitsemuksellisen palvelun tuottaessaan glukoosia hermosoluille. Lisäksi niillä on todennäköisesti rooli ionisessa homeostaasissa solunulkoisen kaliumpitoisuuden ja GABA: n kaltaisten välittäjäaineiden aineenvaihdunnan säätelyssä.
3. Mikrogliasoluja.
kolmannen gliasolutyypin oletetaan olevan mesodermaalista alkuperää, joten ne eivät tarkkaan ottaen ole neurogliaalisia kuten astrosyytit ja Muller-solut. Ne tulevat verkkokalvolle samaan aikaan kehitteillä olevien verkkokalvon verisuonten mesenkymaalisten esiasteiden kanssa (Chan-Ling, 1994). Mikrogliaaliset solut ovat kaikkialla ihmisen verkkokalvossa, joka löytyy jokaisesta verkkokalvon kerroksesta.
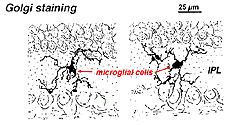
Golgin värjäämässä verkkokalvossa ne näyttävät oudoilta, moninapaisilta muodoilta, joissa on pieniä solukappaleita ja epäsäännöllisiä lyhyitä prosesseja. Golgi-valmisteissa niitä onkin joskus luultu hermosoluiksi varsinkin silloin, kun ne ovat ydinkerroksessa, jonka prosessit ovat yksikäsitteisesti pleksikerroksessa.
kuva. 9.
Mikrogrammasolujen Golgi-värjäystä voi olla kahta tyyppiä. Yhden muodon ajatellaan tulevan verkkokalvolle jo varhaisessa kehitysvaiheessa näköhermon mesenkymestä ja makaavan lepotilassa verkkokalvon kerroksissa suuren osan verkkokalvon eliniästä. Toinen mikroglian muoto näyttää olevan veren välityksellä kulkeutuvia soluja, jotka ovat mahdollisesti peräisin perisyyteistä (Boycott and Hopkins, 1981; Gallego, 1986). Molemmat tyypit voidaan stimuloida makrofagiseen toimintaan verkkokalvon trauman jälkeen, ja sitten ne harjoittavat rappeutuvien verkkokalvon neuronien fagosytoosia.

Fig. 10 a. Lektiinillä värjättyä mikrogliaalista solua Chan-Liniltä vuodelta 1994.

Kuva. 10B. ihmisen verkkokalvo, jossa on astrosyyttejä ja Mikrogliaa. Tämä kuva Scott McLeod Jerry Lutty lab, on koko ihmisen verkkokalvon valmistelu kolminkertainen laneled fluoresoivia vasta-aineita, jotka värjäävät verisuonia (sininen), astrosyytit (punainen) ja mikroglia (vihreä).
4. Viittaus.
Boycott BB, Hopkins JM. Mikroglia verkkokalvossa apinoiden ja muiden nisäkkäiden; sen ero muihin glia-ja vaakasuoriin soluihin. Neurotiede.1981;6:679–688.
Cajal SR. In: Thorpe SA, Glickstein M, translators. 1892. Verkkokalvon rakenne. Springfield (IL): Thomas; 1972.
Chan-Ling T. Glial, neuronaaliset ja vaskulaariset interaktiot nisäkkään verkkokalvolla. Prog. Ret. Eye Res. 1994;13: 357-389.
Edwards RB. Retinoiinihapon biosynteesi Müllerin gliasolujen avulla: malli keskushermostolle? Prog. Ret. Eye Res. 1994;13: 231-242.
Fisher SK, Lewis GP. Photoreceptors and beyond: cellular and molecular effects of verkkokalvon irtauma.2nd Great Basin Visual Science Symposium, II, University of Utah Press. 1995
Frishman LJ, Steinberg RH. Valon aiheuttama o: n lisääntyminen pimeässä sopeutuneen kissan verkkokalvon proksimaalisessa osassa. J Neurofysioli. 1989;61:1233–1243.
Gallego A. comparative studies on horizontal cells and a note on microglial cells. Prog. Ret. 1986; 5: 165-206.
Guerin CJ, Anderson DH, Fisher SK. Välihehkulangan immunolabeloinnissa tapahtuu muutoksia, jotka johtuvat verkkokalvon irtaumasta ja kiinnittymisestä uudelleen kädellisillä.Sijoittaa. Ophthal. Vis. Sci. 1990;31:1474–1482.
Karwoski CJ, Proenza LM. Mullerin solujen vasteiden, transretinaalisen potentiaalin ja kaliumvuon välinen suhde. J Neurofysioli. 1977;40:244–259.
Miller RF, Dowling JE. Mudpuppy retinan Muller-solujen (gliasolujen) solunsisäiset vasteet: niiden suhde elektroretinogrammin B-aaltoon. J Neurofysioli.1970;33:323–341.
Newman EA, Odette LL. Malli elektroretinogrammi B-Aalto sukupolvi: testi K+ hypoteesi. J Neurofysioli. 1984;51:164–182.
Newman EA. Elektrofysiologia verkkokalvon gliasolujen. Prog. Ret. 1989;8:153-172.
Reichenbach A, Robinson SR. Müllerin solujen osallisuus verkkokalvossa. Julkaisussa: Djamgoz MBA, Archer SN, Vallerga s, editors. Neurobiologia ja ulkoisen verkkokalvon kliiniset näkökohdat. Lontoo: Chapman & Hall; 1995. 395-416.
Robinson SR, Hampson ECGM, Munro MN, Vaney DI. Neuroglian välisten rakoliitosten yksisuuntainen kytkentä. Proc. Austr. Neurotieteilijä. Soc. 1993;3:167.
Schnitzer J. astrosyytit nisäkkäiden verkkokalvossa. Prog. Ret. 1988;7:209-232.
Kivi J, Makarov F, Hollander H. verkkokalvon ganglionsolujen Soman ja aksonikukkulan glial ensheathment. Vis Neurosci. 12:273–279.
Kivi J, Dreher Z. astrosyyttien, gangliosolujen ja verkkokalvon verisuoniston välinen suhde. J Comp Neurol. 1987;255:35–49.
trivino a, Ramirez JM, Salazar JJ, Ramirez AI, Garcia-Sanchez J. immunohistokemiallinen tutkimus ihmisen näköhermon pään astrogliasta. Vision Res. 1996;36: 2015-2028.
Turner DL, Cepko CL. Hermosolujen ja glia: n yhteinen kantaisä säilyy rotan verkkokalvolla kehityksen loppupuolella. Luonto. 1987;328:131–136.