pyruvaattikinaasi m2 ruokkii syöpäsolujen useita näkökohtia: solun aineenvaihdunnasta, transkriptiosäätelystä solunulkoiseen signalointiin
biokemiallinen analyysi, jossa määriteltiin entsymaattinen aktiivisuus, joka katalysoi laktaatin muodostumista glukoosista solulysaateissa, paljasti ensimmäisen solunsisäisen metaboliareitin, glykolyyttisen reitin. Glykolyyttistä toimintaa sisältävien fraktioiden puhdistamisesta alkaen joukko pioneeritutkijoita auttoi tunnistamaan entsyymit, jotka osallistuvat reitin jokaiseen vaiheeseen . Nämä tulokset rakentavat modernin konseptimme aerobisen ja anaerobisen hengityksen sekä energiantuotannon vaihdossa erilaisissa fysiologisissa ja patologisissa olosuhteissa.
erään entsyymin olemassaolosta, joka katalysoi ATP: n tuotantoa siirtämällä fosfaattiryhmän Pep: stä ADP: hen maksassa, raportoitiin ensimmäisen kerran vuonna 1934 . Entsyymin myöhempi eristäminen, joka tunnetaan myöhemmin nimellä pyruvaattikinaasi (PK), osoitti eroja kudosjakautumisessa ja katalyyttisessä kinetiikassa, mikä viittaa siihen, että tällä entsyymillä voi olla erilaisia isoformeja . Vuosien 1982-1984 aikana kloonattiin erilaisia PK-geenejä hiivasta, kanasta ja rotasta . Pkm2: n funktionaalinen tutkimus aloitettiin 1980-luvun alussa, kun hiirestä tunnistettiin kandidaattigeeni . Myöhemmin, Noguchi ym. osoitti, että kaksi isoformeja PK (PKM1 ja PKM2) koodataan sama PKM geeni kautta vaihtoehtoista liitos . Ihmisillä myös PKM-isoformeja tuotetaan samankaltaisella liitosmekanismilla sisällyttämällä eksoni 9 ja 10 pkm1: een ja pkm2 mRNA: han erikseen .
useat havainnot kiinnittivät tutkijan huomion pkm2: n mahdolliseen rooliin tuumorigeneesissä. Ensinnäkin pkm2 on alkion isoformi, joka ilmenee voimakkaasti eläinten kehityksen aikana. Sen transkriptio vaimenee useissa aikuisten kudoksissa samalla kun se aktivoituu uudelleen kasvaimissa . Toiseksi pkm1: n ja pkm2: n suhteellisen runsauden tutkimus normaaleissa ja kasvainkudoksissa osoitti siirtymisen PKM1: n isoformista PKM2: n isoformiin erilaisissa syövissä, kuten maksasolusyövässä . Kolmanneksi mRNA: n siirtämistä pkm1: stä pkm2: een parantaa c-Myc-onkogeeni, joka viittaa syöpäsolujen aktiivisesti osallistuvan tähän vaihtoon, jotta ne sopivat vaatimuksiinsa proliferaatiossa ja aineenvaihdunnassa . Neljänneksi pkm2: n aktiivisuuden Mukauttaminen aktivaattoreilla tai inhibiittoreilla vaikuttaa kasvaimen kasvuun in vivo .
- ensimmäinen jakso: PKM2 metabolisena entsyyminä sytoplasmassa
- toinen jakso: pkm2 toimii sytoplasmassa signalointimodulaattorina
- kolmas jakso: PKM2 transkription säätelijänä tumassa
- neljäs jakso: pkm2 solunulkoisena signalointiviestijänä
- The fifth episode: pkm2: n
- jatkuu jakso: onko pkm2 syövän biomarkkeri ja lääkekohde?
ensimmäinen jakso: PKM2 metabolisena entsyyminä sytoplasmassa
koska pkm2: n roolia glykolyysin aineenvaihdunnassa syöpäsoluissa on tarkasteltu laajasti , tiivistämme tässä vain kolme ratkaisevaa eroa PKM1 – ja PKM2-välitteisen katalyysin ja solujen metabolian välillä. Ensimmäinen ero on alayksikön vuorovaikutus. Sekä PKM1 että PKM2 ovat neljän identtisen alayksikön muodostamia tetrameerisiä proteiineja. Jokainen alayksikkö (tai monomeeri) sisältää neljä rakenteellista domeenia, joihin kuuluvat A -, B -, C-ja N-päätteinen domeeni. Monomeeri dimerisoituu ensin yhteen ja sitten kaksi dimeeriä vuorovaikuttaa monomeerin C-domeenin orkestroiman dimeeri-dimeerirajapinnan kautta muodostaen tetrameerin. Koska pkm1 ja PKM2 sisältävät eri eksoneja mrnoissaan, tämä muuttaa C-domeenin koodattuja aminohappoja ja muuttaa tetrameerin stabiilisuutta. Fysiologisessa tilassa PKM1 järjestäytyy konstitutiivisesti tetrameeriksi, kun taas PKM2 voi olla olemassa tetrameerinä tai dimeerinä. Toinen ero on allosteerinen sääntely. Pienten molekyylien ja metaboliittien solunsisäisistä pitoisuuksista riippuen PKM1: n ja PKM2: n aktiivisuutta voidaan säädellä eri tavoin. Yksi tunnetuimmista allosteerisista säätelijöistä on fruktoosi-1,6-bisfosfaatti (FBP). Tämä glykolyyttinen Välituote sitoutuu suoraan PKM2: een ja lisää pkm2: n affiniteettia PEP: hen . Päinvastoin, FBP ei vaikuta merkittävästi PKM1: n toimintaan. FBP: n lisäksi muiden metaboliittien, aminohappojen ja pienten molekyylien on raportoitu vaikuttavan PKM2: n aktiivisuuteen (Fig. 1). Aktivaatioon tai inhibitioon tarvittava pitoisuus on kuitenkin suuri ja moduloiva vaikutus vaatimaton. On vielä epäselvää, onko näillä pienillä molekyyleillä tärkeä rooli PKM: n aktiivisuuden säätelyssä fysiologisissa olosuhteissa. PKM2: n aktiivisuutta säätelevät myös translaation jälkeiset modifikaatiot, kuten fosforylaatio, asetylaatio ja hapettuminen, jotka suosivat dimeerisen PKM2: n vähäistä aktiivisuutta (Kuva. 1). Kolmas ero on energiantuotanto ja välituotekäyttö. Koska pkm1 on konstitutiivisesti olemassa aktiivisena tetrameerinä, tämän isoformin tärkein biologinen tehtävä on ATP: n tuottaminen soluenergian tuottamiseksi. PKM2 voi kuitenkin ATP: n tuottamisen lisäksi siirtyä vähemmän aktiiviseen dimeeriseen muotoon tuottamaan useita glykolyyttisiä välituotteita, joita voidaan käyttää rakennuspaloina aminohappojen, lipidien ja nukleotidien biosynteesissä.
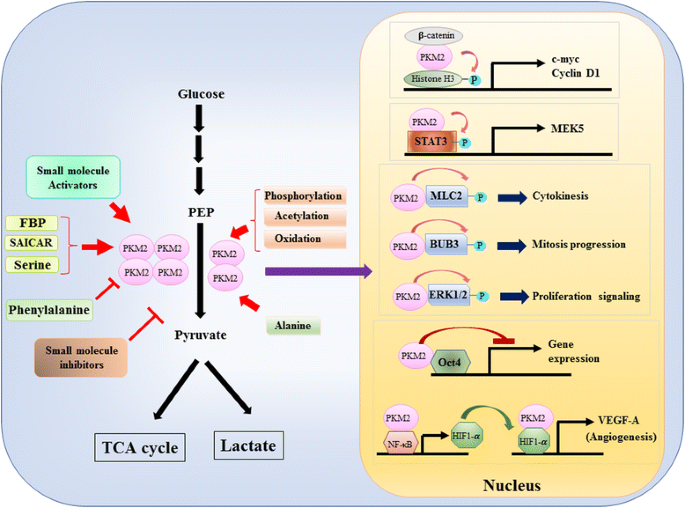
pkm2: n aktiivisuuden Mukauttaminen fysiologisten molekyylien avulla ja translaation jälkeinen modifikaatio ja pkm2: n mahdollinen rooli transkription säätelyssä
toinen jakso: pkm2 toimii sytoplasmassa signalointimodulaattorina
glykolyyttisenä entsyyminä, minkä lisäksi pkm2: n ehdotetaan osallistuvan useampiin soluprosesseihin, koska sytoplasmassa tunnistetaan vuorovaikutuksessa olevia proteiineja. Esimerkiksi PKM2: n osoitettiin olevan vuorovaikutuksessa useiden tyrosiinikinaasien kanssa, mukaan lukien A-Raf, Break point Cluster region-Abelson (BCR-ABL) – fuusiokinaasi, fibroblastin kasvutekijäreseptori 1 (FGFR1) jne. . Näiden sitoutumiskumppanien on osoitettu moduloivan pkm2: n dimeeristä/tetrameeristä muutosta solujen metabolian muuttamiseksi. On kuitenkin mahdollista, että PKM2 voi vastavuoroisesti vaikuttaa näiden sitoutumiskumppaneiden katalyyttiseen kinetiikkaan, substraatin sitoutumiseen ja sytoplasmaan sijaintiin moduloidakseen signaalinsiirtoa. Havainto siitä, että PKM2 on fosforityrosiinia sitova proteiini, vahvistaa tätä mahdollisuutta, koska monet solunsisäiset signalointivälittäjät voivat sitoutua fosforityrosiinijäämiin kootakseen erityisiä proteiinikomplekseja signaalinsiirtoa varten . Tähän mennessä pkm2: n sitovien kumppaneiden lista kasvaa jatkuvasti. Korostamme useita uusia jäseniä ja keskustelemme heidän biologisista vaikutuksistaan täällä. Mukheree ym. on osoitettu, että PKM2 voi sitoutua hur: iin, RNA: ta sitovaan proteiiniin, jolla on tärkeä rooli mRNA: n stabiilisuuden ja translaatiotehokkuuden säätelyssä ja edistää solusyklin etenemistä ja glioomasolujen proliferaatiota . Mielenkiintoista, toinen RNA sitova proteiini tristetraproliini, joka voisi sitoa useita mRNA kautta AU-rikas Elementti 3 ’- kääntämätön Alue (3 ’ – UTR) havaittiin myös olevan pkm2 vuorovaikutuksessa kumppani, ja PKM2 indusoi fosforylaatio ja hajoaminen tristetraproliini moduloida rintasyövän kasvua . Nämä kaksi tutkimusta viittaavat pkm2: n mahdolliseen translationaaliseen kontrollifunktioon. Äskettäin Liang et al. tunnistettu anti-apoptoottinen proteiini Bcl2 uudeksi pkm2 kumppaniksi . He osoittivat, että oksidatiivinen stressi aiheutti pkm2: n translokaation mitokondrioihin, joissa se fosforyloi ja stabiloi Bcl2: ta estämällä sen hajoamisen ubikitinaatiosta riippuvaisen reitin kautta. Nämä tiedot viittasivat siihen, että pkm2 auttaa syöpäsoluja sopeutumaan oksidatiiviseen stressiin, jonka aiheuttaa solunsisäinen metabolinen muutos tai solunulkoinen loukkaus.
kolmas jakso: PKM2 transkription säätelijänä tumassa
pkm2: n ydinroolia geenin transkription tai epigeneettisen modifikaation säätelyssä ehdotti ensinnäkin havainto, että PKM2 sitoutui Y333-fosforoituneeseen β-kateniiniin ja β-kateniini-PKM2-kompleksi värvättiin nukleosomeihin fosforyloimaan Histoni H3 treoniini 11: ssä (kuva. 1). Tämä fosforylaatio lisäsi myöhemmin Histoni H3-asetylaatiota, joka johti β-kateniinin kohdegeenien säätelyyn. Toinen pkm2: n suoraan fosforyloima transkriptiotekijä on transkription 3 (STAT3) signaalianturi ja aktivaattori . PKM2-välitteinen STAT3: n fosforylaatio tyrosiini 705: ssä lisäsi STAT3: n aktiivisuutta mitogeeniaktivoidun proteiinikinaasi 5: n (MEK5) ilmentymisen säätelyssä. Transkriptiotekijöiden lisäksi PKM2: n on osoitettu fosforyloivan myosiinin valoketjua 2 (MLC2), BUB3: A ja solunulkoista signaalia säätelevää kinaasia 1 ja 2 (ERK1 ja ERK2) . Mielenkiintoista, PKM2 toimii myös kautta fosforylaatio-riippumaton tavalla vaikuttaa geenin ilmentymistä. Esimerkiksi PKM2: n on havaittu sitoutuvan Oct4: ään, joka on yksi kantasolujen itsensä uusiutumista ohjaavista master-transkriptiotekijöistä ja estää Oct4-välitteistä transkriptiota . PKM2 voi myös tehostaa kasvaimen angiogeneesiä vuorovaikutuksessa NF-kB: n ja HIF-1α: n kanssa tumassa ja aktivoida HIF-1α: n kohdegeenin VEGF-A: n ilmentymisen .VEGF-A: n lisääntynyt eritys tehostaa verisuonten muodostumista, mikä edistää kasvaimen kasvua. Vaikka nämä tutkimukset viittasivat vahvasti pkm2: n ydinlokalisoitumiseen ja proteiinikinaasin toimintaan erilaisissa fysiologisissa ja patologisissa olosuhteissa, pkm2-välitteisen geeniekspression merkitys on kuitenkin kyseenalaistettu PKM2-tyrmäyssoluja käyttävissä tutkimuksissa. Käyttämällä-merkitty PEP ja PKM2 – null hiiren alkion fibroblastit, Hosios et al. osoitti, että PEP-riippuvainen fosforylaatio ei ole yleinen tapahtuma soluissa eikä reaktiota katalysoi PKM2 . Näiden tutkimusten ristiriita on tällä hetkellä ratkaisematta, ja pkm2: n proteiinikinaasiaktiivisuus tarvitsee lisävahvistusta.
neljäs jakso: pkm2 solunulkoisena signalointiviestijänä
solunulkoisen PKM2: n esiintyminen avaa uuden väylän PKM2: n biologisen toiminnan tutkimiselle. Buschow ym. toimitti ensimmäiset todisteet siitä, että PKM2: ta voitiin havaita B-solujen eksosomeissa ja että se tunnistettiin MHC-luokkaan II kuuluvaksi proteiiniksi . Kaksi myöhempää tutkimusta osoitti myös, että PKM2: ta esiintyy eri solujen vapauttamissa eksosomeissa . Tällä hetkellä useat julkiset tietokannat, kuten ExoCarta ja EVpedia, tarjoavat kattavaa tietoa eri lajien solunulkoisten rakkuloiden sisältämistä komponenteista, kuten proteiineista, lipideistä ja nukleiinihapoista. Kaikki tiedot vahvistavat, että PKM2 on eksosomien pakettiproteiini. Viimeaikaiset tutkimukset ovat selvästi osoittaneet eksosomien kommunikatiivisen roolin toimittamalla eri komponentteja isäntäsoluista vastaanottajasoluihin . On odotettavissa, että PKM2 voi olla rooli solu-solu ylikuulumisen.
esiin tulevat todisteet todellakin tukevat tätä hypoteesia. Esimerkiksi tuore tutkimus osoitti, että pkm2: ta kiertävä veri voi edistää kasvaimen kasvua ja angiogeneesiä lisäämällä endoteelisolujen kasvua, muuttoliikettä ja matriisi-adheesiota . Toinen tutkimus osoitti myös, että paksusuolisyöpäsoluista erittyvä PKM2 saattaa vaikuttaa itsevaltiaan stimulaation kautta solujen migraation tehostamiseksi aktivoimalla PI3K/Akt-ja Wnt/β-kateniinireitit . Syöpäsolujen lisäksi kudosvauriokohtien neutrofiilit voivat vapauttaa pkm2: ta angiogeneesin ja haavan paranemisen edistämiseksi . Tuore tutkimuksemme osoitti myös, että rekombinantti PKM2-proteiini voi aiheuttaa fosforylaatiota ja epidermaalisen kasvutekijäreseptorin (EGFR) aktivoitumista . Lisäksi havaitsimme, että r339e-mutantti PKM2, joka ensisijaisesti muodosti dimeerisen PKM2: n, aktivoi EGFR: ää merkittävämmin kuin tetramerinen PKM2. Keller ym. tunnistettiin 154 proteiinia mahdollisiksi pkm2: n substraateiksi, kun Hela-soluja on käsitelty sukkinyyli-5-aminoimidatsoli-4-karboksamidi-1-riboosi-5′ – fosfaatilla (SAICAR), joka on solunsisäinen metaboliitti, joka voi stimuloida pkm2: n proteiinikinaasiaktiivisuutta . He löysivät myös EGFR: n pkm2-substraattina. Heidän tuloksensa poikkeavat meidän tuloksistamme kahdella tavalla. Ensinnäkin tutkimuksessamme aktivoidut signalointireitit saavat alkunsa solunulkoisesta pkm2: sta, kun taas heidän tutkimuksessaan tunnistetut molekyylikohteet ovat mahdollisia solunsisäisen PKM2: n substraatteja. Toiseksi erk1/2-aktiivisuuden lisääntyminen tutkimuksessamme aloitetaan EGFR-aktivaatiolla, kun taas ERK1/2-aktivaatiota heidän tutkimuksessaan stimuloi suoraan SAICAR/PKM2-kompleksi. Yksi samanlainen ilmiö havaittu molemmissa tutkimuksissa on, että R339E mutantti PKM2 aktivoi signalointimolekyylejä merkittävämmin kuin villi tyyppi PKM2 viittaa erillinen rooli dimeerinen ja tetrameerinen PKM2 onkogeneesissä. Käyttämällä reseptorityrosiinikinaasi array, havaitsimme, että solunulkoinen PKM2 aktivoi vain rajoitettu kasvutekijän reseptorit rintasyöpäsolujen (tiedot eivät näy). Tällä hetkellä solunulkoisen pkm2: n reseptoriaktivaation selektiivisyyttä ei tunneta. Lisäksi ei ole selvää, miksi R339E-mutantti PKM2 on voimakkaampi EGFR: n aktivaatiossa. Näihin kysymyksiin vastaamiseksi tarvitaan lisää kokeiluja.
toinen tyylikäs kysymys on, onko vapaalla PKM2: lla ja vesikkelipakkauksella pkm2: lla samanlainen vaikutus tumorigeneesin edistämisessä (Kuva. 2). Vapaa solunulkoinen pkm2 ei voinut tunkeutua plasman kalvoon ja se pystyi aktivoimaan solunsisäistä signalointia vain solun pintaproteiinien, kuten kasvutekijäreseptorien kautta. Vastaavasti vesikkeli-Pakkaus PKM2 voisi olla endosytoitu syöpä-ja stroomasoluissa, ja vesikkeleistä vapautuva pkm2 voisi vaikuttaa aineenvaihduntaan ja geeniekspressioon solunsisäisen mekanismin kautta. The PKM2-null cells or mice will be useful to elucidate whether these two extracellular forms of PKM2 could act synergistically or antagonistically in tumorigenesis.

The potential tumor-promoting effect of extracellular PKM2
The fifth episode: pkm2: n
vastaamaton ristiriita edellä käsiteltyjen solupohjaisten tietojen lisäksi myös pkm2: n onkogeeninen rooli on kyseenalaistettu PKM2 knockout-hiirten sukupolven jälkeen. Israelsen ym. loi ehdollisen knockout-hiirimallin poistamalla pkm2-spesifisen eksonin 10 . Yllättäen pkm2: n ehtyminen nopeutti, mutta ei heikentänyt kasvaimen muodostumista, jonka taustalla oli BRCA1-geenin menetys hiirillä. Nämä tiedot osoittivat, että PKM2: ta ei tarvita syöpäsolujen proliferaatioon. Mielenkiintoista, PKM1 expression havaittiin vain ei-proliferoituvissa kasvainsoluissa viittaa kasvain-suppressiivinen rooli pkm1 rintasyövässä. Lisäksi PKM2 knockout hiirillä on suuri esiintyvyys kehittää hepatosellulaarinen karsinooma spontaanisti pitkän latenssin jälkeen johtuen aineenvaihdunnan epätasapainosta . Nämä tulokset viittaavat siihen, että PKM2: lla on onkogeeninen rooli in vivo.
jatkuu jakso: onko pkm2 syövän biomarkkeri ja lääkekohde?
vaikka geenitekniikan hiirimallin tulokset eivät tue pkm2: n kasvainta edistävää aktiivisuutta, pkm2: n yliekspressiota esiintyy yleisesti ihmisen syövissä ja se liittyy huonoon kliiniseen tulokseen (Taulukko 1) . Myös kaksi tuoretta meta-analyysitutkimusta tukivat tätä päätelmää. Wu ym. analysoitiin tiedot 2812 potilaiden kiinteitä kasvaimia ruoansulatuskanavan saatu 16 kohortti tutkimukset ja todettiin, että yli-ilmentymä PKM2 liittyy alentunut kokonaiselossaolo mahasyöpä, ruokatorven okasolusyöpä, hepatosellulaarinen karsinooma, sappisyöpä ja suusyöpä . Pkm2 ei kuitenkaan ole haimasyövän ennustava tekijä. Tämä havainto ei ole yhdenmukainen aiempien tutkimusten kanssa, joiden mukaan pkm2: n lisääntyminen on osoitus haimasyöpäpotilaiden huonosta eloonjäämisestä . Toiseen tutkimukseen sisältyi 4796 tapausta 27 yksittäisestä tutkimuksesta, jotka osoittivat yhdistetyissä tiedoissa, että PKM2: n säätelyn lisääntyminen korreloi huonompaan kokonaiselinaikaan, taudista vapaaseen elossaoloon ja uusiutumisesta vapaaseen elossaoloon . Syöpätyypin mukaan kerrostettuna pkm2 ei kuitenkaan ennusta haimasyövän heikkoa selviytymistä. Kollektiivisesti, PKM2 näyttää olevan vastuussa ennusteen markkeri useimmissa kiinteissä kasvaimissa.
sen sijaan pkm2: n käyttö diagnostisena tekijänä on kiistanalaista. Proteomianalyysi osoitti, että PKM2 on mahdollinen diagnostinen markkeri keuhkosyövän toteamisessa . Tuoreen tutkimuksen mukaan PKM2 ei kuitenkaan ole hyvä keuhkosyövän diagnostinen merkkiaine alhaisen spesifisyyden vuoksi . Vastaavasti pelkästään pkm2 ei todennäköisesti ole hyödyllinen merkkiaine paksusuolen syövän seulonnassa . Useiden merkkiaineiden yhdistäminen voisi kuitenkin lisätä herkkyyttä ja spesifisyyttä syöpädiagnoosissa .
pkm2: n terapeuttinen potentiaali on kiehtova tapahtuma syövän hoidossa. Yhdeltä puolelta pkm2: n eston odotetaan estävän glykolyysiä, heikentävän geenien transkriptiota ja estävän solujen proliferaatiota. Siksi PKM2-estäjät näyttävät olevan hyviä ehdokkaita syöpälääkkeiden kehittämiseen. Käyttämällä kirjaston seulonta, Vander Heiden et al. tunnistettiin kolme uutta luokkaa PKM2: n estäjiä ja osoitettiin, että tehokkain yhdiste esti pkm2: n toimintaa ja aiheutti syöpäsolujen kuoleman . Äskettäin Ning et al. todettiin, että uudet naftokinonijohdannaiset ovat voimakkaita PKM2: n estäjiä . Yksi tehokas yhdiste 3 k tukahdutti useiden syöpäsolulinjojen proliferaation alle mikromolaarisina pitoisuuksina, vaikka se ei juurikaan vaikuttanut haitallisesti normaaleihin soluihin. Toiselta puolelta, aktivointi PKM2 voi myös estää kasvaimen kasvua. Koska alhainen aktiivisuus PKM2 dimeeri on tärkein isoformi, joka laukaisee glykolyysiä sytoplasmassa ja geenitranskriptio tumassa syöpäsoluissa, pkm2-aktivaattorit, jotka voivat edistää tetrameerisen PKM2: n muodostumista, voivat vaihtaa glykolyysiä mitokondrioiden reitille ja vähentää ydinvoiman tuloa heikentämään geenin transkriptiota. Molemmat vaikutukset heikentävät aineenvaihdunnan kysyntää ja kasvua tukevaa signalointia, joka johtaa kasvaimen regressioon. Kahdessa pioneeritutkimuksessa tunnistettiin erilaisia PKM2-aktivaattoreita ja luonnehdittiin niiden spesifisyyttä in vitro . Myöhempi tutkimus osoitti, että PKM2-aktivaattorit todella edistivät tetrameerin muodostumista ja tukahduttivat kasvaimen kasvua In vivo . Nämä tulokset viittasivat siihen, että PKM2-aktivaattorit voisivat olla lupaavia syöpälääkkeitä.
Solunsalpaajaresistenssi on syövän hoidossa merkittävä tukos. Syöpäsolujen luontaisen kemoresistenssin voittaminen on kiireellistä kliinistä tutkimusta varten. Pkm2: n roolit syöpäsolujen kemoresistenssissä on paljastettu, ja pkm2: n kohdentamisen on osoitettu herkistävän uudelleen kemoresistenttejä syöpäsoluja. Tuore tutkimus osoitti, että CD44 on vuorovaikutuksessa PKM2: n kanssa ja estää PKM2: n toimintaa lisäämällä Pkm2: n Tyr105-fosforylaatiota. CD44-ablaatio aiheutti siirtymisen aerobisesta glykolyysistä mitokondriohengitykseen ja lisääntyneeseen reaktiivisten happilajien (Ros) tuotantoon, mikä johti sisplatiinin herkkyyden lisääntymiseen kolorektaalisyöpäsoluissa . PKM2: n aktiivisuuden eston on osoitettu estävän glykolyysiä ja voittavan sisplatiiniresistenssin. Siksi sisplatiinin ja PKM2: n estäjien yhdistelmä voi olla tehokas strategia kemoterapiassa (Fig. 3). PKM2: n on myös raportoitu osallistuvan gemsitabiiniresistenssin säätelyyn haimasyöpäsoluissa. Kim ym. osoitti, että PKM2 edistää gemsitabiiniresistenssiä säätelemällä negatiivisesti P38-välitteistä p53-fosforylaatiota, mikä vähentää p53: n transkriptioaktiivisuutta ja estää pro-apoptoottisten geenien ilmentymisen . Lisäksi r399e-PKM2: n kohdunulkoinen ilmentyminen, joka muodostaa ensisijaisesti dimeeristä PKM2: ta, parantaa haimasyövän solujen vastustuskykyä gemsitabiinille. Nämä todisteet viittaavat siihen, että PKM2 edistää lääkeresistenssin induktiota ei-metabolisen mekanismin kautta. Koska dimeerinen PKM2 toimii pääasiassa transcriptionaalisena säätelijänä syöpäsoluissa, pkm2-aktivaattorit, jotka edistävät tetrameerisen PKM2: n muodostumista, voivat tehokkaasti tukahduttaa pkm2: n ei-metabolisen toiminnan ja vähentää gemsitabiiniresistenssiä haimasyöpäsoluissa. Pkm2: n ydinvoiman on myös osoitettu edistävän EGFR-estäjien vastustuskykyä kolorektaalisyövässä ja keuhkosyövässä . Li ym. osoitti, että pkm2-välitteinen STAT3-fosforylaatio vähentää kolorektaalisyövän solujen herkkyyttä gefitinibille ja että pkm2: n ja STAT3: n palautetun gefitinibiherkkyyden vuorovaikutus soluissa häiriintyy . Äskettäin toinen tutkimus osoitti, että PKM2 translokoituu tumaan ja vuorovaikuttaa poly-ADP-riboosin (PAR) kanssa kasvutekijän stimulaation yhteydessä. PKM2: n PAR-sitoutumisaktiivisuus on kriittinen pkm2: n ydinsäilytyksen ja geenitranskription kannalta ja tärkeä solujen proliferaation ja kasvaimen kasvun edistämisen kannalta. He päättelivät, että pkm2: n ydintoiminnan estäminen voi voittaa EGFR-mutatoituneiden syöpäsolujen resistenssin . Molemmat tutkimukset osoittivat, että pkm2: lla on keskeinen rooli lääkeresistenssin välittäjänä syövissä, ja ehdottivat, että pkm2: n kohdistaminen ydinenergiaan voi olla lupaava strategia vastarinnan kumoamiseksi.

PKM2 edistää proliferaatiota ja lääkeaineresistenssiä syöpäsoluissa