Piruvato chinasi M2 combustibili molteplici aspetti di cellule di cancro: dal metabolismo cellulare, la regolazione trascrizionale di segnalazione extracellulare
analisi Biochimiche da caratterizzante l’attività enzimatica che catalizza la formazione di lattato dal glucosio in lisati cellulari rivelato il primo intracellulare via metabolica, pathway glicolitico. A partire dalla purificazione delle frazioni che contenevano attività glicolitica, un certo numero di ricercatori pionieri ha contribuito all’identificazione di enzimi che coinvolgono in ogni fase del percorso . Questi risultati costruiscono il nostro concetto moderno nello scambio di respirazione aerobica e anaerobica e produzione di energia in varie circostanze fisiologiche e patologiche.
L’esistenza di un enzima che catalizzava la produzione di ATP trasferendo un gruppo fosfato dal PEP all’ADP nel fegato fu riportata per la prima volta nel 1934 . Il successivo isolamento dell’enzima, noto in seguito come piruvato chinasi (PK), ha dimostrato differenze nella distribuzione tissutale e nella cinetica catalitica suggerendo che questo enzima può avere isoforme diverse . Durante 1982 a 1984, vari geni PK sono stati clonati da lievito, pollo e ratto . Lo studio funzionale di PKM2 è stato avviato con l’identificazione di un gene candidato nel topo nei primi anni 1980 . Più tardi, Noguchi et al. ha dimostrato che due isoforme di PK (PKM1 e PKM2) sono codificate dallo stesso gene PKM tramite splicing alternativo . Nell’uomo, le isoforme di PKM inoltre sono prodotte via un simile meccanismo d’impionbatura includendo esclusivamente l’esone 9 e 10 in mRNA PKM1 e PKM2 .
Diversi risultati hanno attirato l’attenzione del ricercatore sul potenziale ruolo della PKM2 nella tumorigenesi. In primo luogo, PKM2 è l’isoforma embrionale altamente espressa durante lo sviluppo animale. La sua trascrizione è attenuata in un certo numero di tessuti adulti mentre viene riattivata nei tumori . In secondo luogo, lo studio dell’abbondanza relativa di PKM1 e PKM2 nei tessuti normali e tumorali ha dimostrato un passaggio dall’isoforma PKM1 all’isoforma PKM2 in vari tumori come il carcinoma epatocellulare . In terzo luogo, il cambiamento di mRNA splicing da PKM1 a PKM2 è migliorato da c-Myc oncogene suggerendo le cellule tumorali attivamente impegnarsi in questo interruttore per soddisfare il loro requisito in proliferazione e metabolismo . In quarto luogo, la modulazione dell’attività PKM2 da parte di attivatori o inibitori influisce sulla crescita del tumore in vivo .
- Il primo episodio: PKM2 come enzima metabolico nel citoplasma
- Il secondo episodio: PKM2 come una segnalazione modulatore nel citoplasma
- Il terzo episodio: PKM2 come regolatore trascrizionale nel nucleo
- Il quarto episodio: PKM2 come comunicatore di segnalazione extracellulare
- The fifth episode: discrepanza senza risposta di PKM2
- L’episodio che continua: PKM2 è un biomarcatore del cancro e un bersaglio di droga?
Il primo episodio: PKM2 come enzima metabolico nel citoplasma
Poiché il ruolo di PKM2 nel controllo metabolico della glicolisi nelle cellule tumorali è stato ampiamente rivisto , riassumiamo solo tre differenze cruciali tra la catalisi mediata da PKM1 e PKM2 e il metabolismo cellulare qui. La prima differenza è l’interazione delle subunità. Sia PKM1 che PKM2 sono proteine tetrameriche formate da quattro subunità identiche. Ogni subunità (o monomero) contiene quattro domini strutturali tra cui A, B, C e dominio N-terminale. Il monomero prima dimerizza insieme e poi due dimeri interagiscono tramite l’interfaccia dimero-dimero orchestrata dal dominio C del monomero per formare un tetramero. Poiché PKM1 e PKM2 includono esoni diversi nei loro MRNA, questo cambia gli amminoacidi codificati nel dominio C e altera la stabilità del tetramero. In condizioni fisiologiche, PKM1 organizza costitutivamente come tetramero mentre PKM2 può essere esistito in tetramero o dimero. La seconda differenza è la regolazione allosterica. A seconda delle concentrazioni intracellulari di piccole molecole e metaboliti, l’attività di PKM1 e PKM2 può essere regolata in modo differenziato. Uno dei regolatori allosterici più noti è il fruttosio-1,6-bisfosfato (FBP). Questo intermedio glicolitico lega direttamente PKM2 e aumenta l’affinità di PKM2 per PEP . Al contrario, l’FBP non influisce in modo significativo sull’attività di PKM1. Oltre a FBP, altri metaboliti, aminoacidi e piccole molecole sono stati segnalati per influenzare l’attività PKM2 (Fig. 1). Tuttavia, la concentrazione richiesta per l’attivazione o l’inibizione è elevata e l’effetto modulatorio è modesto. Non è ancora chiaro se queste piccole molecole svolgano un ruolo importante nel controllo dell’attività del PKM in circostanze fisiologiche. L’attività di PKM2 è anche regolata dalla modifica post-traslazionale, come la fosforilazione, l’acetilazione e l’ossidazione, che favoriscono la bassa attività del PKM2 dimerico (Fig. 1). La terza differenza è la produzione di energia e l’utilizzo intermedio. Poiché PKM1 esiste costitutivamente come tetramero attivo, la principale funzione biologica di questa isoforma è la generazione di ATP per fornire energia cellulare. Tuttavia, PKM2, oltre a produrre ATP, può passare alla forma dimerica meno attiva per generare diversi intermedi glicolitici che possono essere utilizzati come elementi costitutivi per la biosintesi di aminoacidi, lipidi e nucleotidi.
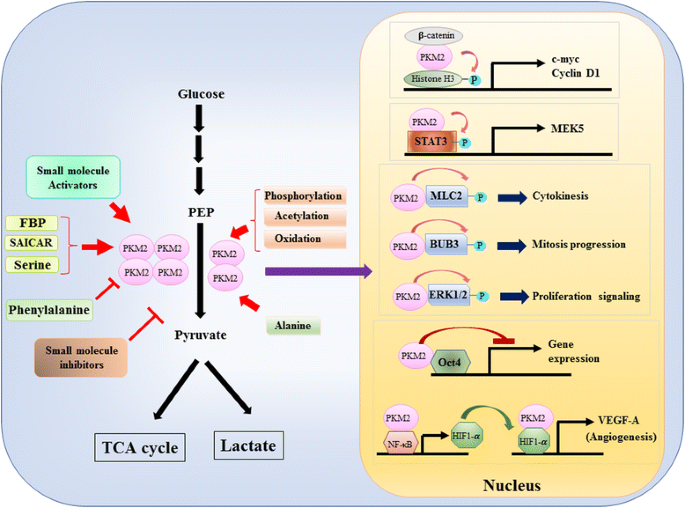
Modulazione di PKM2 attività fisiologiche molecole e modifica alberino-di traduzione e il ruolo potenziale di PKM2 nella regolazione trascrizionale
Il secondo episodio: PKM2 come una segnalazione modulatore nel citoplasma
oltre a funzionare come un enzima glicolitico, PKM2 è proposto di coinvolgere di più i processi cellulari grazie per l’identificazione di proteine interagenti nel citoplasma. Ad esempio, PKM2 ha dimostrato di essere una proteina interagente di diverse tirosin chinasi tra cui A-Raf, Break point cluster region-Abelson (BCR-ABL) fusion chinasi, recettore del fattore di crescita dei fibroblasti 1 (FGFR1) ecc. . Questi partner di legame hanno dimostrato di modulare il cambiamento dimerico/tetramerico di PKM2 per alterare il metabolismo cellulare. Tuttavia, è possibile che PKM2 possa influenzare reciprocamente la cinetica catalitica, il legame del substrato e la posizione citoplasmatica di questi partner di legame per modulare la trasduzione del segnale. La scoperta che PKM2 è una proteina legante fosforo-tirosina rafforza questa possibilità perché molti mediatori di segnalazione intracellulari possono legarsi ai residui di fosforo-tirosina per assemblare complessi proteici specifici per la trasmissione del segnale . Ad oggi, l’elenco dei partner vincolanti PKM2 cresce continuamente. Evidenziamo diversi nuovi membri e discutiamo la loro implicazione biologica qui. Mukheriee et al. dimostrato che PKM2 potrebbe legarsi con HuR, una proteina legante RNA che svolge un ruolo importante nel controllo della stabilità dell’mRNA e dell’efficienza traslazionale, per promuovere la progressione del ciclo cellulare e la proliferazione delle cellule di glioma . È interessante notare che un’altra proteina di legame dell’RNA tristetraprolin che potrebbe legare un certo numero di mRNA attraverso l’elemento ricco di AU nella regione 3′-untranslational (3′-UTR) è stata trovata anche come un partner interagente PKM2, e la fosforilazione indotta da PKM2 e la degradazione della tristetraprolina per modulare la crescita del cancro al seno . Questi due studi implicano una potenziale funzione di controllo traslazionale di PKM2. Recentemente, Liang et al. identificato la proteina anti-apoptotica Bcl2 come nuovo partner PKM2 . Hanno dimostrato che lo stress ossidativo ha indotto la traslocazione di PKM2 nei mitocondri dove ha fosforilato e stabilizzato Bcl2 prevenendo la sua degradazione attraverso la via ubiquitinazione-dipendente. Questi dati hanno suggerito che PKM2 aiuta le cellule tumorali ad adattare lo stress ossidativo provocato dal cambiamento metabolico intracellulare o dall’insulto extracellulare.
Il terzo episodio: PKM2 come regolatore trascrizionale nel nucleo
Un ruolo nucleare di PKM2 nella regolazione della trascrizione genica o della modificazione epigenetica è stato inizialmente suggerito dalla scoperta che PKM2 si legava con β-catenina fosforilata Y333 e il complesso β-catenina-PKM2 è stato reclutato nei nucleosomi per fosforilare l’istone H3 a treonina 11 (Fig. 1). Questa fosforilazione ha successivamente aumentato l’acetilazione dell’istone H3 che ha portato alla sovraregolazione dei geni bersaglio della β-catenina. Un altro fattore di trascrizione direttamente fosforilato da PKM2 è trasduttore di segnale e attivatore della trascrizione 3 (STAT3) . La fosforilazione mediata da PKM2 di STAT3 a tirosina 705 ha migliorato l’attività di STAT3 per upregulate l’espressione della chinasi proteica attivata dal mitogeno chinasi 5 (MEK5). Accanto ai fattori di trascrizione, PKM2 ha dimostrato di fosforilare la catena leggera della miosina 2 (MLC2), BUB3 e chinasi 1 e 2 regolate dal segnale extracellulare (ERK1 ed ERK2). È interessante notare che PKM2 agisce anche tramite modalità indipendente dalla fosforilazione per influenzare l’espressione genica. Ad esempio, PKM2 è stato trovato per legarsi con Oct4, uno dei fattori di trascrizione master che controllano l’auto-rinnovamento delle cellule staminali e inibiscono la trascrizione mediata da Oct4 . PKM2 può anche migliorare l’angiogenesi tumorale interagendo con NF-kB e HIF-1α nel nucleo e attivando l’espressione del gene bersaglio HIF-1α VEGF-A. Di conseguenza, l’aumento della secrezione di VEGF-A aumenta la formazione dei vasi sanguigni che contribuisce alla crescita del tumore . Sebbene questi studi suggerissero fortemente la localizzazione nucleare e la funzione della proteina chinasi di PKM2 in varie circostanze fisiologiche e patologiche, tuttavia l’importanza dell’espressione genica mediata da PKM2 nucleare è stata messa in discussione da studi che utilizzano cellule knockout PKM2. Utilizzando-marcato PEP e PKM2-null mouse fibroblasti embrionali, Hosios et al. ha dimostrato che la fosforilazione PEP-dipendente non è un evento comune nelle cellule e la reazione non è catalizzata da PKM2 . La discrepanza di questi studi è attualmente irrisolta e l’attività della protein chinasi di PKM2 ha bisogno di ulteriore conferma.
Il quarto episodio: PKM2 come comunicatore di segnalazione extracellulare
La presenza di PKM2 extracellulare apre una nuova strada per lo studio della funzione biologica di PKM2. Buschow et al. ha fornito la prima prova che PKM2 potrebbe essere rilevato negli esosomi delle cellule B ed è stato identificato come una proteina associata a MHC di classe II . Due studi successivi hanno anche indicato che PKM2 esiste in esosomi rilasciati da varie cellule . Attualmente, diversi database pubblici come ExoCarta e EVpedia forniscono informazioni complete per i componenti tra cui proteine, lipidi, acidi nucleici di vescicole extracellulari in diverse specie. Tutti i dati confermano che PKM2 è una proteina del pacchetto degli esosomi. Studi recenti hanno chiaramente dimostrato un ruolo comunicativo degli esosomi fornendo diversi componenti dalle cellule ospiti alle cellule riceventi . È prevedibile che PKM2 possa svolgere un ruolo nella diafonia cellulare-cellulare.
Le prove emergenti supportano effettivamente questa ipotesi. Ad esempio, un recente studio ha dimostrato che il sangue circolante PKM2 può promuovere la crescita del tumore e l’angiogenesi aumentando la crescita, la migrazione e l’adesione della matrice delle cellule endoteliali . Un’altra indagine ha anche dimostrato che PKM2 secreto dalle cellule tumorali del colon potrebbe agire tramite una stimolazione autocrina per migliorare la migrazione cellulare attivando le vie PI3K / Akt e Wnt / β-catenina . Oltre alle cellule tumorali, i neutrofili nei siti di danno dei tessuti potrebbero rilasciare PKM2 per promuovere l’angiogenesi e la guarigione delle ferite . Il nostro recente studio ha anche dimostrato che la proteina PKM2 ricombinante potrebbe indurre la fosforilazione e l’attivazione del recettore del fattore di crescita epidermico (EGFR) . Inoltre, abbiamo scoperto che il mutante PKM2 R339E che formava preferenzialmente PKM2 dimerico attivava l’EGFR in modo più significativo rispetto al tetramerico PKM2. Keller et al. identificato 154 proteine come potenziali substrati per PKM2 dopo il trattamento delle cellule Hela con succinil-5-amminoimidazolo-4-carbossamide-1-ribosio-5′-fosfato (SAICAR), un metabolita intracellulare che potrebbe stimolare l’attività della protein chinasi di PKM2 . Hanno anche trovato EGFR come substrato PKM2. I loro risultati sono diversi dai nostri in due modi. Innanzitutto, le vie di segnalazione attivate nel nostro studio sono provocate da PKM2 extracellulare mentre i bersagli molecolari identificati nel loro studio sono potenziali substrati di PKM2 intracellulare. In secondo luogo, l’aumento dell’attività di ERK1/2 nel nostro studio è iniziato dall’attivazione di EGFR mentre l’attivazione di ERK1/2 nel loro studio è direttamente stimolata dal complesso SAICAR/PKM2. Un fenomeno simile osservato in entrambi gli studi è che il mutante R339E PKM2 attiva le molecole di segnalazione in modo più significativo rispetto al tipo selvaggio PKM2 suggerendo il ruolo distinto del PKM2 dimerico e tetramerico nell’oncogenesi. Utilizzando il recettore tirosina chinasi array, abbiamo scoperto che extracellulare PKM2 attivato solo recettori del fattore di crescita limitato nelle cellule di cancro al seno (dati non mostrati). Attualmente, la selettività dell’attivazione del recettore da parte di PKM2 extracellulare rimane sconosciuta. Inoltre, il motivo per cui il mutante R339E PKM2 è più potente nell’attivazione di EGFR non è chiaro. Sono necessari ulteriori esperimenti per rispondere a queste domande.
Un’altra domanda elegante da affrontare è se il PKM2 libero e il PKM2 per l’imballaggio delle vescicole mostrano un effetto simile nel promuovere la tumorigenesi (Fig. 2). Il PKM2 extracellulare libero non poteva penetrare la membrana del plasma e poteva soltanto attivare la segnalazione intracellulare via le proteine di superficie delle cellule come i ricevitori di fattore di crescita. Al contrario, PKM2 di vescicola-imballaggio potrebbe essere endocytosed dalle cellule del cancro e dello stroma, e il PKM2 rilasciato dalle vescicole potrebbe influenzare il metabolismo e l’espressione genica tramite meccanismo intracellulare. The PKM2-null cells or mice will be useful to elucidate whether these two extracellular forms of PKM2 could act synergistically or antagonistically in tumorigenesis.

The potential tumor-promoting effect of extracellular PKM2
The fifth episode: discrepanza senza risposta di PKM2
Oltre ai dati basati sulle cellule discussi sopra, il ruolo oncogeno di PKM2 è stato messo in discussione anche dopo la generazione di topi knockout PKM2. Israelsen et al. generato un modello di mouse knockout condizionale eliminando l’esone 10 specifico per PKM2 . Sorprendentemente, l’esaurimento di PKM2 ha accelerato ma non attenuato la formazione del tumore guidata dalla perdita del gene Brca1 nei topi. Questi dati hanno indicato che PKM2 non è richiesto per la proliferazione delle cellule tumorali. È interessante notare che l’espressione di PKM1 è stata rilevata solo nelle cellule tumorali non proliferanti, suggerendo un ruolo soppressivo del tumore di PKM1 nel cancro al seno. Inoltre, i topi knockout PKM2 hanno un’alta incidenza di sviluppare spontaneamente il carcinoma epatocellulare dopo una lunga latenza a causa dello squilibrio nel metabolismo . Questi risultati contro la nozione che PKM2 svolge un ruolo oncogenico in vivo.
L’episodio che continua: PKM2 è un biomarcatore del cancro e un bersaglio di droga?
Sebbene i risultati del modello murino geneticamente ingegnerizzato non supportino l’attività di promozione del tumore di PKM2, la sovraespressione di PKM2 è universalmente riscontrata nei tumori umani ed è associata a scarsi risultati clinici (Tabella 1) . Due recenti studi di meta-analisi hanno anche supportato questa conclusione. Wu et al. analizzato i dati di 2812 pazienti con tumori solidi del sistema digestivo ottenuti da 16 studi di coorte e ha scoperto che la sovraespressione di PKM2 è associata a una ridotta sopravvivenza globale nel cancro gastrico, carcinoma a cellule squamose esofagee, carcinoma epatocellulare, cancro biliare e cancro orale . Tuttavia, PKM2 non è un fattore prognostico per il cancro del pancreas. Questa scoperta non è coerente con studi precedenti che dimostrano che l’aumento di PKM2 è un indicatore di scarsa sopravvivenza dei pazienti con cancro al pancreas . Un’altra indagine ha incluso 4796 casi provenienti da 27 studi individuali che hanno dimostrato che la sovraregolazione della PKM2 è correlata con una sopravvivenza globale peggiore, una sopravvivenza libera da malattia e una sopravvivenza libera da recidive in dati aggregati . Tuttavia, stratificato per tipo di cancro, PKM2 non prevede una scarsa sopravvivenza del cancro al pancreas. Collettivamente, PKM2 sembra essere un marcatore prognostico responsabile nella maggior parte dei tumori solidi.
al contrario, l’uso di PKM2 come fattore diagnostico è controversa. Un’analisi proteomica ha dimostrato che PKM2 è un potenziale marcatore diagnostico per la rilevazione del cancro del polmone . Tuttavia, uno studio recente ha suggerito che PKM2 non è un buon marcatore diagnostico per il cancro del polmone a causa della bassa specificità . Allo stesso modo, PKM2 da solo è improbabile che sia un marcatore utile per lo screening del cancro del colon . Tuttavia, la combinazione di marcatori multipli potrebbe aumentare la sensibilità e la specificità per la diagnosi del cancro .
Il potenziale terapeutico di PKM2 è un evento intrigante nel trattamento del cancro. Da un lato, si prevede che l’inibizione di PKM2 inibisca la glicolisi, comprometta la trascrizione genica e sopprima la proliferazione cellulare. Pertanto, gli inibitori di PKM2 sembrano essere buoni candidati per lo sviluppo di farmaci anti-cancro. Usando lo screening della biblioteca, Vander Heiden et al. ha identificato tre nuove classi di inibitori di PKM2 e ha dimostrato che il composto più efficace inibiva l’attività di PKM2 e induceva la morte delle cellule tumorali . Recentemente, Ning et al. ha scoperto che i nuovi derivati del naftochinone sono potenti inibitori della PKM2 . Un efficace composto 3 k ha soppresso la proliferazione di più linee cellulari tumorali a concentrazioni sub-micromolari mentre ha mostrato poco effetto dannoso sulle cellule normali. Dall’altro lato, l’attivazione di PKM2 può anche inibire la crescita del tumore. Poiché il dimero PKM2 a bassa attività è l’isoforma principale che innesca la glicolisi nel citoplasma e la trascrizione genica nel nucleo nelle cellule tumorali, gli attivatori PKM2 che possono promuovere la formazione di PKM2 tetramerico possono passare la glicolisi alla via dei mitocondri e ridurre l’ingresso nucleare per attenuare la trascrizione genica. Entrambi gli effetti compromettono la domanda metabolica e la segnalazione di supporto alla crescita che porta alla regressione del tumore. Due studi pionieristici hanno identificato vari attivatori di PKM2 e ne hanno caratterizzato la specificità in vitro . Uno studio successivo ha dimostrato che gli attivatori di PKM2 hanno effettivamente promosso la formazione del tetramero e hanno soppresso la crescita tumorale in vivo . Questi risultati hanno suggerito che gli attivatori di PKM2 potrebbero essere promettenti farmaci anti-cancro.
La resistenza alla chemioterapia è un blocco importante per il trattamento del cancro. Il superamento della chemioresistenza intrinseca delle cellule tumorali è urgente per la ricerca clinica. I ruoli di PKM2 nella chemoresistenza delle cellule tumorali sono stati rivelati e il targeting di PKM2 ha dimostrato di ri-sensibilizzare le cellule tumorali chemoresistenti. Uno studio recente ha indicato che CD44 interagisce con PKM2 e sopprime l’attività PKM2 via la fosforilazione aumentante di Tyr105 di PKM2. L’ablazione CD44 ha indotto il passaggio dalla glicolisi aerobica alla respirazione mitocondriale e all’aumento della produzione di specie reattive dell’ossigeno (ROS), con conseguente aumento della sensibilità al cisplatino nelle cellule tumorali del colon-retto . L’inibizione dell’attività PKM2 è stata dimostrata per sopprimere la glicolisi e superare la resistenza al cisplatino. Pertanto, la combinazione di cisplatino e inibitori della PKM2 può essere una strategia efficace per la chemioterapia (Fig. 3). PKM2 è stato anche segnalato per partecipare alla regolazione della resistenza alla gemcitabina nelle cellule tumorali pancreatiche. Kim et al. ha dimostrato che PKM2 promuove la resistenza alla gemcitabina attraverso la regolazione negativa della fosforilazione p53 mediata da p38, riducendo quindi l’attività trascrizionale di p53 e sopprimendo l’espressione dei geni pro-apoptotici . Inoltre, l’espressione ectopica di R399E-PKM2, che forma preferenzialmente PKM2 dimerico, aumenta la resistenza delle cellule tumorali pancreatiche alla gemcitabina. Queste evidenze suggeriscono che PKM2 contribuisce all’induzione della resistenza ai farmaci attraverso un meccanismo non metabolico. Poiché il PKM2 dimerico agisce principalmente come regolatore trascrizionale nelle cellule tumorali, gli attivatori PKM2 che promuovono la formazione di PKM2 tetramerico possono essere efficaci nel sopprimere la funzione non metabolica di PKM2 e ridurre la resistenza alla gemcitabina nelle cellule tumorali pancreatiche. Il PKM2 nucleare inoltre è stato dimostrato per contribuire alla resistenza dell’inibitore di EGFR nel cancro colorettale e nel cancro polmonare . Li et al. ha dimostrato che la fosforilazione STAT3 mediata da PKM2 nucleare riduce la sensibilità delle cellule tumorali del colon-retto a gefitinib e l’interruzione dell’interazione tra PKM2 nucleare e STAT3 ha ripristinato la sensibilità gefitinib nelle cellule . Più recentemente, un altro studio ha dimostrato che PKM2 si trasferisce nel nucleo e interagisce con il ribosio poli-ADP (PAR) sulla stimolazione del fattore di crescita. L’attività PAR-legante di PKM2 è critica per conservazione nucleare e la trascrizione del gene di PKM2 ed è importante per la promozione della proliferazione delle cellule e della crescita del tumore. Hanno concluso che l’inibizione della funzione nucleare di PKM2 può superare la resistenza delle cellule tumorali mutate da EGFR . Entrambi gli studi hanno sottolineato un ruolo cruciale del PKM2 nucleare nel mediare la resistenza ai farmaci nei tumori e hanno suggerito che il targeting del PKM2 nucleare potrebbe essere una strategia promettente per ignorare la resistenza.
 figure3
figure3PKM2 contribuisce alla proliferazione e alla resistenza ai farmaci nelle cellule tumorali