Uno zucchero speciale: come l’acido sialico influisce sul metabolismo, sulla salute e sulle malattie
Stiamo brulicando di microbi che vivono su superfici esterne e interne al nostro corpo. La nostra comprensione del microbioma intestinale, in particolare, sta rapidamente migliorando quando iniziamo a scoprire come complesse comunità microbiche interagiscono con la nostra dieta, con altri microbi e con le nostre superfici cellulari. Mentre molte sostanze chimiche sono importanti in questo ambiente, qui ci concentriamo sugli acidi sialici come un insieme critico di molecole che sono alla base di molte di queste interazioni e impatto sul metabolismo, sulla salute e sulle malattie.
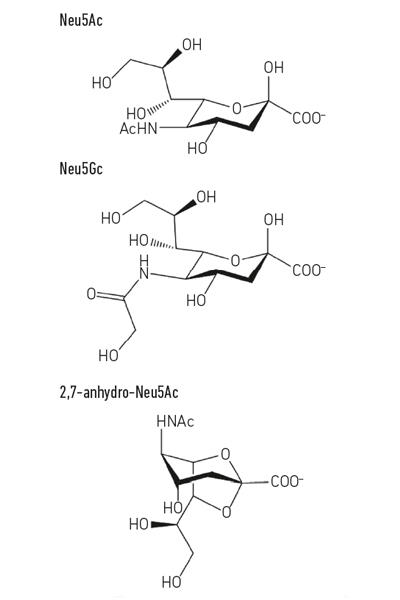
L’acido sialico più comune, l’acido N-acetil-neuraminico (Neu5Ac) (Fig. 1) è un acido zuccherino relativamente semplice che, a parte la sua carica, non ha nulla di intrinsecamente insolito. Tuttavia, quando le cellule contattano altre cellule usano spesso molecole visualizzate sulla superficie, ed è qui che gli acidi sialici diventano così importanti, poiché di solito sono gli zuccheri terminali sui glicani ospiti che pepano proteine e lipidi seduti sulle superfici cellulari. Le glicoproteine della mucina secrete o della cellula-superficie sono buoni esempi delle strutture acido-rivestite sialic che i microbi potrebbero incontrare alle superfici mucose, quale il tratto respiratorio o gastrointestinale (GI). Questa posizione unica come molecola “meet and greet” per altre cellule umane, cellule batteriche e virus mette gli acidi sialici al centro della scena per molti processi importanti.
- Fig. 1. Esempio di derivati dell’acido sialico. Si noti che l’acido N-glicolil-neurnaminico (Neu5Gc) non può essere prodotto dall’uomo, una differenza biochimica che ci distingue dalle altre grandi scimmie, ma può essere incorporato nelle nostre superfici cellulari quando acquisito dalla dieta.
- Agenti patogeni ricoperti di zucchero
- Cross-feeding – condividere i dolci intorno
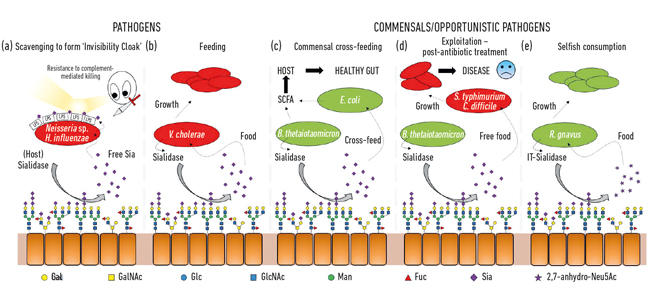
- Fig. 2. Illustrazione a fumetti delle varie interazioni tra batteri e acidi sialici presenti come zuccheri terminali su una gamma di glicani ospiti. Questi sono scissi dalle sialidasi e di solito convertiti in Neu5Ac libero con l’eccezione della IT-sialidasi che rilascia 2,7-anidro-Neu5Ac. L’acido sialico libero rilasciato dall’ospite o dalle sialidasi patogene può quindi essere utilizzato dai patogeni come mantello di invisibilità (a) o per l’alimentazione (b). L’alimentazione incrociata può verificarsi tra membri del microbiota intestinale o tra batteri commensali e patogeni che condividono la stessa nicchia (c, d) mentre altri batteri hanno sviluppato meccanismi egoistici per riservare l’acido sialico per il proprio consumo (e).
- Tenere in mano i propri dolci
- I virus entrano in azione
- Ulteriori letture
- Andrew Bell
- Emanuele Severi
- Nathalie Juge
- Gavin Thomas H.
- Perché la microbiologia è importante?
- Che consiglio daresti a qualcuno che inizia in questo campo?
Fig. 1. Esempio di derivati dell’acido sialico. Si noti che l’acido N-glicolil-neurnaminico (Neu5Gc) non può essere prodotto dall’uomo, una differenza biochimica che ci distingue dalle altre grandi scimmie, ma può essere incorporato nelle nostre superfici cellulari quando acquisito dalla dieta.
Agenti patogeni ricoperti di zucchero
Una delle prime funzioni microbiche degli acidi sialici è stata collegata alla capacità di alcuni agenti patogeni di colonizzare e quindi causare malattie nell’uomo. Infatti, l’ex presidente della Microbiology Society, il professor Harry Smith FRS, lavorando presso l’Università di Birmingham, ha aperto la strada a questo lavoro con Neisseria gonorrea. Quando gli scienziati del suo gruppo hanno isolato l’insetto direttamente dal corpo, era resistente all’uccisione da parte del siero umano (uccisione mediata dal complemento), ma se gli insetti sono stati coltivati in laboratorio per alcune generazioni, questa protezione è stata persa e le cellule sono state uccise dal siero. Il suo gruppo aveva scoperto che questi agenti patogeni rubano l’acido sialico dall’ospite e lo usano per rivestire la propria superficie cellulare per umanizzarli. Tuttavia, poiché possono solo pulire questo acido sialico, la sua assenza nei terreni di crescita significa che non viene aggiunto ulteriore acido sialico, quindi perdono rapidamente questa “protezione del siero” e vengono uccisi. Si scopre che una serie di altri insetti subdoli usano strategie simili per generare il proprio mantello di invisibilità usando acido sialico, come Neisseria meningitidis, Campylobacter jejuni o Haemophilus influenzae (Fig. 2a)
Cross-feeding – condividere i dolci intorno
Oltre ad essere utilizzati dai patogeni come un mantello di invisibilità, gli acidi sialici sono piuttosto buoni alimenti per i batteri, in quanto forniscono sia carbonio che azoto e possono essere utilizzati come fonte di energia. Lavori precedenti hanno mostrato che agenti patogeni come Vibrio cholerae erano felici solo di mangiare acidi sialici e che di per sé era importante per una colonizzazione ospite di successo (Fig. 2 ter).
Nell’intestino, una fonte importante di acidi sialici proviene dalle mucine, che sono i principali componenti strutturali dello strato di muco che copre la superficie dell’epitelio. Lo strato di muco più esposto è l’habitat di vari batteri commensali, che hanno imparato a banchettare con le proteine della mucina, dove gli zuccheri costituiscono l ‘ 80% della loro massa. Da una semplice prospettiva topologica, gli acidi sialici, come gli zuccheri terminali, vengono raccolti per primi, esponendo gli zuccheri sottostanti che possono anche essere successivamente rilasciati e mangiati. Con l’avvento del sequenziamento di nuova generazione, abbiamo acquisito conoscenze sui microbi che vivono in questa nicchia e molti hanno geni per l’assorbimento dell’acido sialico e il successivo catabolismo. Ciò che è stato emozionante è stata la scoperta che alcuni degli anaerobi commensali, come Bacteroides thetaiotaomicron, secernono sialidasi per rilasciare l’acido sialico, ma mancano di trasportatori e geni catabolici per usarlo effettivamente. Questo gesto apparentemente altruistico è richiesto ai batteri di accedere agli zuccheri sottostanti e sostiene un processo di cross-alimentazione metabolica ad altri insetti che vivono nella nicchia del muco, come i ceppi di Escherichia coli commensali che non hanno una sialidasi ma sono bravi a mangiare acido sialico libero (Fig. 2 quater). Questo concetto è ora riconosciuto come un aspetto importante della colonizzazione e dell’adattamento di nicchia. Una brillante esemplificazione di questo è stata descritta dal gruppo di Justin Sonnenburg della Stanford University, utilizzando un modello murino con un semplice microbiota intestinale definito. Dopo il trattamento antibiotico, molti dei batteri che mangiano acido sialico sono stati spazzati via, con conseguente accumulo di acido sialico libero nell’intestino. Quando poi sfidati con agenti patogeni come Salmonella typhimurium o Clostridium difficile, questi agenti patogeni trarrebbero beneficio dall’acido sialico libero come nutriente (Fig. 2d). I ceppi mutanti di agenti patogeni privi dei trasportatori o dei geni catabolici non hanno avuto un vantaggio, creando una connessione diretta alla crescita del patogeno quando la comunità microbica intestinale viene interrotta. Sono stati quindi proposti meccanismi per ridurre il pool di acido sialico libero nell’intestino come parte di una nuova strategia anti – infettiva.
Fig. 2. Illustrazione a fumetti delle varie interazioni tra batteri e acidi sialici presenti come zuccheri terminali su una gamma di glicani ospiti. Questi sono scissi dalle sialidasi e di solito convertiti in Neu5Ac libero con l’eccezione della IT-sialidasi che rilascia 2,7-anidro-Neu5Ac. L’acido sialico libero rilasciato dall’ospite o dalle sialidasi patogene può quindi essere utilizzato dai patogeni come mantello di invisibilità (a) o per l’alimentazione (b). L’alimentazione incrociata può verificarsi tra membri del microbiota intestinale o tra batteri commensali e patogeni che condividono la stessa nicchia (c, d) mentre altri batteri hanno sviluppato meccanismi egoistici per riservare l’acido sialico per il proprio consumo (e).
Tenere in mano i propri dolci
Mentre la sialomicrobiologia molto precoce si è concentrata sugli agenti patogeni, il lavoro più recente si è concentrato sui batteri commensali che abitano la stessa nicchia. Gli studi di oggi nel campo del microbiota intestinale stanno rivelando quanto sia feroce la competizione per Neu5Ac nel nostro intestino e come possa far pendere l’equilibrio tra patogeni e commensali. E se un particolare batterio potesse rilasciare acido sialico in una forma a cui solo lui potrebbe accedere? Il lavoro nel gruppo Juge a Norwich ha mostrato che il simbionte intestinale Ruminococcus gnavus si è evoluto per farlo, utilizzando un tipo speciale di sialidasi chiamata trans-sialidasi intramolecolare (IT-sialidasi) che rilascia 2,7-anidro-Neu5Ac (vedi Fig. 1) invece di Neu5Ac. Sviluppando un metodo per sintetizzare 2,7-anidro-Neu5Ac puro, hanno dimostrato che i ceppi di R. gnavus erano in grado di crescere su 2,7-anidro-Neu5Ac come unica fonte di carbonio. Oltre al dominio che catalizza la reazione enzimatica, le IT-sialidasi comprendono un dominio di legame dei carboidrati che aiuta a mediare il legame della IT-sialidasi alle mucine ricche di acido sialico. In vivo, ciò può favorire un meccanismo che aiuta a indirizzare i batteri verso le regioni ricche di acido sialico del tratto gastrointestinale, promuovendo quindi la colonizzazione batterica all’interno dello strato di muco. Una volta situato nel posto giusto, l’IT-sialidasi scinde l’acido sialico dalle catene terminali delle mucine e lo rilascia in una forma, 2,7-anidro-Neu5Ac, che R. gnavus può utilizzare come propria fonte di cibo, piuttosto che condividere Neu5Ac con altri batteri che abitano la nicchia del muco, compresi gli agenti patogeni (Fig. 2e). Questo meccanismo fornisce un vantaggio competitivo rispetto ad altri batteri che mangiano acido sialico che abitano la nicchia del muco, fornendo una fonte di nutrienti a cui può accedere preferenzialmente. Inoltre, R. gnavus può anche aiutare a ridurre la crescita di agenti patogeni enterici riducendo il livello di Neu5Ac nell’ambiente mucoso, aprendo la strada a una nuova strategia terapeutica in un’epoca di crescente resistenza agli antibiotici.
I virus entrano in azione
I virus usano anche gli acidi sialici come molecole chiave durante il loro ciclo di infezione, il più famoso virus influenzale. L’attaccamento iniziale è il primo passo chiave in quanto gli acidi sialici funzionano come recettori per il virus sulle cellule epiteliali respiratorie. Il tipo di legame dell’acido sialico con gli zuccheri sottostanti nei glicani controlla spesso la gamma ospite dei virus influenzali e i ceppi pandemici hanno spesso proteine di adesione (emoagglutinina) che riconoscono le forme simili all’uomo di attaccamento all’acido sialico e quindi sono particolarmente virulenti. L’acido sialico è importante nell’ultima fase del ciclo di vita virale poiché il rilascio dalla cellula infetta richiede la scissione degli acidi sialici dalla superficie della cellula ospite. Questo è catalizzato da un enzima virale chiamato sialidasi (o neuraminidasi). L’inibizione di questo enzima da parte di analoghi strutturali dell’acido sialico, formulati in Tamiflu® (Oseltamivir phosphate) e Zanamivir, sono il nostro trattamento di prima linea per l’influenza e sono un trionfo della struttura/funzione ispirata al design del farmaco.
In conclusione, è chiaro che gli acidi sialici sono molecole critiche in molti processi in cui cellule microbiche e virus interagiscono con le superfici cellulari dei mammiferi. In questo breve articolo, abbiamo toccato alcuni di questi per dare un’idea delle funzioni centrali di questi zuccheri nella comunicazione, stealth, colonizzazione e metabolismo in diversi aspetti della brughiera e della malattia. Gli acidi sialici sono stati descritti come le sostanze chimiche più importanti al mondo e senza dubbio continueranno a rivelare di più sull’evoluzione degli esseri umani e sulla loro interazione con i loro microbi.
Ulteriori letture
Thomas, GH. Acquisizione di acido sialico nei batteri – un substrato, molti trasportatori. Biochem Soc Trans 2016;44: 760-765.
Ng KM, Ferreyra JA, Higginbottom SK, Lynch JB, Kashyap PC et al. Gli zuccheri ospiti liberati dal microbiota facilitano l’espansione post-antibiotica dei patogeni enterici. Natura 2013;502: 96-99.
Owen CD, Tailford LE, Monaco S, Šuligoj T, Vaux L et al. Svelare la specificità e il meccanismo di riconoscimento dell’acido sialico da parte del simbionte intestinale Ruminococcus gnavus. Nat Commun 2017; 8: 2196.
Tailford LE, Owen CD, Walshaw J, Crost EH, Hardy-Goddard J, Le Gall G, de Vos WM, Taylor GL, Juge N. La scoperta delle trans-sialidasi intramolecolari nel microbiota intestinale umano suggerisce nuovi meccanismi di adattamento della mucosa. Nat Commun 2015; 6: 7624.
Juge N, Tailford L, Owen CD. Sialidasi da batteri intestinali: una mini-recensione. Biochem Soc Trans 2016;44:166-175.

Andrew Bell
Quadram Institute Bioscience, Norwich Research Park, NR4 7UQ, UK
Andrew Bell è uno studente post-laurea nel Dipartimento Gut Microbes and Health dell’Istituto Quadram che studia l’impatto di un nuovo meccanismo del metabolismo dell’acido sialico sull’omeostasi intestinale. Ha ottenuto il suo BSc in biochimica presso l’Università di East Anglia e ha trascorso 2 anni di lavoro presso il John Innes Centre sul metabolismo della clorofilla in pea (Pisum sativum) prima di entrare nel gruppo di Nathalie Juge al Quadram Institute nel 2015 per il suo lavoro di dottorato.

Emanuele Severi
Dipartimento di Biologia, Università di York, Wentworth Modo, York YO10 5DD, UK
@emm_severi
Emanuele Severi è un post-dottorato di ricerca associato presso il Dipartimento di Biologia dell’Università di York, lavorando nel laboratorio del Prof Gavin Thomas. Ha lavorato sul trasporto e sul metabolismo dell’acido sialico in diversi momenti della sua carriera ed è membro della Società dal 2002.

Nathalie Juge
Quadram Istituto di Bioscienze, Norwich Research Park, NR4 7UQ, UK
Nathalie Juge è un Leader di Ricerca presso il Quadram Istituto di Bioscienze (QIB) in Norwich, Vice dirigente dell’Istituto Programma Strategico, i Microbi intestinali e la Salute (GMH), e Professore Onorario presso la Scuola di Scienze Biologiche, Università di East Anglia. Dirige un gruppo di ricerca sulla glicobiologia delle interazioni ospite-microbo nell’intestino con particolare attenzione al metabolismo dell’acido sialico derivato dalla mucina da parte del microbiota intestinale (https://quadram.ac.uk/nathalie-juge).

Gavin Thomas H.
Dipartimento di Biologia, Università di York, Wentworth Modo, York YO10 5DD, UK
@GavinHThomas
Gavin H. Thomas è un Prof. di Microbiologia del Dipartimento di Biologia dell’Università di York e hanno lavorato sui batteri di acido sialico trasporto e catabolismo per oltre un decennio. È stato membro della Società dal 1995, è stato redattore di Microbiology Today (2003-2006), ha fatto parte del Consiglio nello stesso periodo, ha fatto parte del comitato per le comunicazioni (2012-2015) e del comitato per la diversità di uguaglianza & (2013-2015). È stato redattore, Senior Editor ed è ora Vice redattore capo della rivista leader della Società Microbiology (http://thomaslabyork.weebly.com).
Perché la microbiologia è importante?
Gavin: È importante per la nostra salute – solo ora stiamo davvero iniziando a capire la composizione molecolare e la funzione del nostro microbioma, che probabilmente influisce sul nostro corpo in molti modi.
Che consiglio daresti a qualcuno che inizia in questo campo?
Gavin: Leggi ampiamente e oltre ‘il tuo bug’ – i microbi possono fare praticamente tutto e di solito hanno inventato diversi modi per farlo!
Immagini: particelle di virus dell’influenza H3N2. CDC / Scienza Fototeca.
Fig. 1. Andrew Bell.
Fig. 2. Emmanuele Severi.