Pathophysiology/International Osteoporosisしょう症財団
ホルモンはおそらく骨形成の最も重要な調節因子である。 Oestrogen Lindsay,R.,osteoporosisしょう症の予防および治療ということは十分に確立されている。 ランセット、1993年。 341(8848):p.801-5.
。 副甲状腺ホルモン、P.、ビタミンD生理。 Prog Biophys Mol Biol、2006。 92(1):p.4-8
. そしてoestrogenへの転換によってより少し程度のテストステロンに直接または間接的にSeeman、E.の人の骨のもろさの構造基礎。 ボーン、1999年。 25(1):p.143-7.
ヴァン-ポッテルベルグI.、ら。、特発性骨粗鬆症とその息子の男性における摂動性ステロイド状態。 J Clin内分泌Metab、2004。 89(10):p.4949-53.
。 最適の骨の開発および維持のために必要です。 これらのうち、エストロゲンは現在、骨芽細胞および破骨細胞の表面上の特定のタンパク質または受容体と相互作用する骨細胞に最も直接的な効果を有すると考えられているZallone、A.、骨芽細胞および破骨細胞に対する直接的および間接的なエストロゲン作用。 2006年にAcad Sciに移籍した。 1068:p.173-9
.
この相互作用は、細胞内のイベントの複雑なチェーンをオフに設定し、同時に骨芽細胞-破骨細胞の通信を妨害しながら、骨芽細胞活エストロゲンの効果は、エストロゲン受容体α(ERa)と呼ばれる特定のタイプの細胞表面受容体を介して媒介され、受容体-ホルモン複合体が特定の遺伝子をオンにするスイッチとして作用する細胞の核にホルモンを結合して輸送する。 ERA受容体は、骨芽細胞の表面上に見出され、エストロゲン受容体関連受容体α(Erra)は、骨細胞の調節において補助的な役割を果たし得る、Bonnelye,E.and J.E.Aubin,Estrogen receptor−related receptor alpha:a mediator o f estrogen response in bone. J Clin Endocrinol Metab,2005. 90(5):p.3115-21.
。 最近の研究はまた、細胞へのエストロゲンの侵入を容易にする性ホルモン結合グロブリン(SHBG)も支持的な役割を果たし得ることを示唆しているGoderie-Plomp,H.W.,et al.、内因性性ホルモン、性ホルモン結合グロブリン、および高齢者の男性および女性における椎骨骨折のリスク:ロッテルダム研究。 J Clin内分泌Metab、2004。 89(7):p.3261-9.
。もちろん、エストロゲンは骨からある程度離れたところで作られ、血流に分泌され、子宮や乳房などの他の組織にも大きな影響を与えます。 しかし、骨生理学に大きな影響を与える他の、局所的に産生されるシグナル伝達分子がある。

Figure taken from Ferrari & Roux, 2019 Pocket Reference to Osteoporosis, S. Ferrari, Roux, C., Editor 2019, Springer International Publishing.
.
Prostaglandins, particularly prostaglandin E2 (PGE2), stimulate both resorption and formation of bone Pilbeam, C.C., J.R. Harrison, and L.G. Raisz, Chapter 54 – Prostaglandins and Bone Metabolism, in Principles of Bone Biology (Second Edition), J.P. Bilezikian, L.G. Raisz, and G.A. Rodan, Editors. 2002, Academic Press: San Diego. p. 979-994
. PGE2は、アラキドン酸と呼ばれる前駆体から様々な骨細胞に形成される脂質である。 PGE2合成の最初のステップは、シクロオキシゲナーゼ2(COX2)と呼ばれる酵素によって行われ、この酵素の阻害剤は、in vivoでの機械的負荷による骨形成の誘導を媒介する動物の機械的ストレスに応答して骨形成を防止することができる。 1996年、J. 11(11):p.1688-93.
。 PGE2は、運動誘発性骨形成のために必要とされることがあります。
COX-2Carboneを阻害する非ステロイド性抗炎症薬を服用している人では、骨折のリスクが増加するという証拠があります。,骨ミネラル密度と非ステロイド性抗炎症薬とアスピリンの使用との関連:シクロオキシゲナーゼ選択性の影響. 2003年にJ-Waveで再放送された。 18(10):p.1795-802
また、増加する可能性があります。 骨の改造を調節するように見える脂質分子の別のセットは、ロイコトリエンです。 また、アラキドン酸に由来し、これらはマウスの骨密度を低下させることが見出されているTraianedes,K.,et al.、5-リポキシゲナーゼ代謝産物は、in vitroで骨形成を阻害する。 1998年、東京農業大学農学部農学科を卒業。 139(7):p.3178-84.
。
これらのホルモンのいずれかが骨の改造にどのように影響するかは、破骨細胞および/または骨芽細胞の活性をどのように変化させるかに依存する。
特定の細胞表面受容体は、骨細胞外からの信号を細胞核に伝達するのに役立ち、細胞活性を調節する異なる遺伝子をオンまたはオフに切り替えるこ これらには、骨形成の強力な誘導因子であるタンパク質のファミリーである骨形態形成タンパク質(Bmp)の受容体が含まれる。
BMP受容体は、骨芽細胞前駆細胞の表面に見出されているMbalaviele,G.,et al.、Β-カテニンおよびBMP-2は、骨芽細胞の分化および新しい骨形成を促進するために相乗作用する。 J細胞バイオケム、2005年。 94(2):p.403-18.
。 Wnt受容体である低密度リポタンパク質(LDL)関連タンパク質5受容体(LRP5)と呼ばれる別の細胞表面受容体も、動物におけるLRP5の喪失が重度の骨粗鬆症、LDL受容体関連タンパク質5(LRP5)は、骨の発生と眼の発達に影響を与えます。 セル、2001年。 107(4):p.513-23.
。 BMPの受容器およびLRP5は行為に骨芽細胞を刺激するために協力するかもしれませんがこれが起こるかもしれない丁度いかに明白にされませんでした。
Sost遺伝子の産物であり、骨細胞によって発現されるSclerostinは、骨芽細胞上のLRP5/6受容体に結合し、Wntシグナル伝達を阻害し、骨形成Bonewald、l.F.、驚くべき骨細胞 2011年、J-Waveで放送された。 26(2):p.229-38.
Li,X.,et al.、スクレロスチンはLRP5/6に結合し、標準的なWntシグナル伝達に拮抗する。 2005年にJ-B-ケムに移籍。 280(20):p.19883-7.
。 副甲状腺ホルモン(PTH)および機械的負荷は、oesteocytes Bellidoによるスクレロスチンの分泌を減少させる、T.、et al。、マウスにおける副甲状腺ホルモンの慢性上昇は、骨細胞によるスクレロスチンの発現を減少させる:骨芽細胞形成のホルモン制御のための新規な機2005年、””を発表した。 146(11):p.4577-83.
Robling,A.G.,et al.、In vivoでの骨の機械的刺激は、Sost/sclerostinの骨細胞発現を減少させる。 2008年、J-Popレーベルより発売された。 283(9):p.5866-75.
。 骨強度に対する強力な特性を有する潜在的な薬物として,sclerostinに対する抗体が開発されている。 Read more on anabolics as treatments.

Figure taken from Ferrari & Roux, 2019 Pocket Reference to Osteoporosis, S. Ferrari, Roux, C., Editor 2019, Springer International Publishing.
. RANK(Nf Κ Bの受容体活性化剤)と呼ばれる細胞表面受容体は、RANKが同族のパートナー RANKリガンド(RANKL)によって活性化されると、破骨細胞前駆細胞を完全に分化させた破骨細胞に発達させる。 破骨細胞分化因子は、オステオプロテゲリン/破骨細胞形成抑制因子のリガンドであり、TRANCE/RANKLと同一である。 1998年、日本大学理工学部物理学科卒業。 95(7):p.3597-602.
Lacey,D.L.,et al.、Osteoprotegerinリガンドは、破骨細胞の分化および活性化を調節するサイトカインである。 セル、1998年。 93(2):p.165-76.
。
RANKLは、実際には、骨芽細胞によって産生され、おそらく多くのシグナル伝達分子の一つであり、骨芽細胞と破骨細胞との間のクロストークを促進し、骨 2002年、”月刊コロコロコミック”に掲載。 20:p.795-823.
。 Osteoprotegerin,osteoblastsによって放出される別のタンパク質Suda,T.,et al.、腫瘍壊死因子受容体およびリガンドファミリーの新しいメンバーによる破骨細胞の分化および機能の調節。 1999年、日本コロムビアより発売された。 20(3):p.345-57.
、RANKLに結合することもでき、RANKとRANKLが接触するのを防ぐためのおとりとして機能します。 RANKL/osteoprotegerinのバランスはosteoporosisしょう症で重大かもしれません。 実際、動物実験では、オステオプロテゲリンの産生の増加が骨量の増加につながることが示され、タンパク質の損失は骨粗鬆症および骨折の増加につ、osteoprotegerin欠損マウスは、早期発症骨粗鬆症と動脈石灰化を開発しています。 1998年、””を発表。 12(9):p.1260-8.
。 RANKLの阻害剤はまた、ヒトにおける骨粗鬆症の潜在的な治療法としての約束を示している。
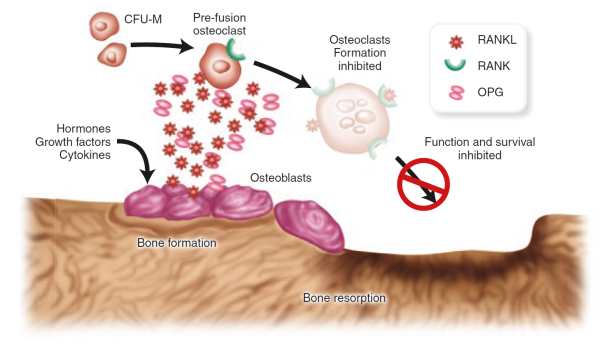
Figure taken from Ferrari & Roux, 2019 Pocket Reference to Osteoporosis, S. Ferrari, Roux, C., Editor 2019, Springer International Publishing.
. 破骨細胞の形成と活性化を促進する第二の相補的な細胞シグナル伝達システムも、ここ数年の間に発見された。
DNAX活性化タンパク質12(DAP12)とFc受容体共通γ鎖(FcRy)、二つの細胞表面受容体の非存在下では、マウスは、骨密度の劇的な増加によって特徴付けられる重度の骨粗しょう症-osteoporosisしょう症の正反対–を開発Mocsai,A.,et al.、免疫調節アダプタータンパク質DAP12とFc受容体γ鎖(FcRgamma)は、Sykチロシンキナーゼを介して機能的破骨細胞の開発を調節します。 2004年、Acad Sci U S Aを設立。 101(16):p.6158-63.
古賀,T.,et al.、ITAMモチーフによって媒介される共刺激信号は、骨の恒常性のためにRANKLと協力する。 ネイチャー、2004年。 428(6984):p.758-63.
。 これら二つの細胞表面受容体は、細胞内カルシウムの増加を引き起こすためにITAM(immunoreceptor tyrosine-based activation motif)アダプタータンパク質と呼ばれる細胞内のタンパク質群と相互作用する。研究は、RANK/RANKLおよびITAM媒介経路が完全な破骨細胞活性を誘導するために協力したことを示唆している。
これら二つの経路は、活性化T細胞の核因子(NFAT)c1と呼ばれるタンパク質を活性化するために収束する可能性があります。 NFATC1は、破骨細胞前駆細胞が完全に活性な破骨細胞になるために必要な遺伝子をオンにするため、骨吸収のマスタースイッチとして機能します高柳,H.,mechanistic insight into osteoclast differentiation in osteoimmunology. JモルMed(Berl),2005. 83(3):p.170-9.
。