Et spesielt sukker: hvordan sialinsyre påvirker metabolisme, helse og sykdom
vi vrimler av mikrober som lever på overflater utenfor og inne i kroppen vår. Vår forståelse av tarmmikrobiomet spesielt forbedrer seg raskt når vi begynner å unpick hvordan komplekse mikrobielle samfunn interagerer med kostholdet vårt, med andre mikrober og med våre celleoverflater. Mens mange kjemikalier er viktige i dette miljøet, fokuserer vi på sialinsyrer som et kritisk sett med molekyler som underbygger mange av disse interaksjonene og påvirker metabolisme, helse og sykdom.
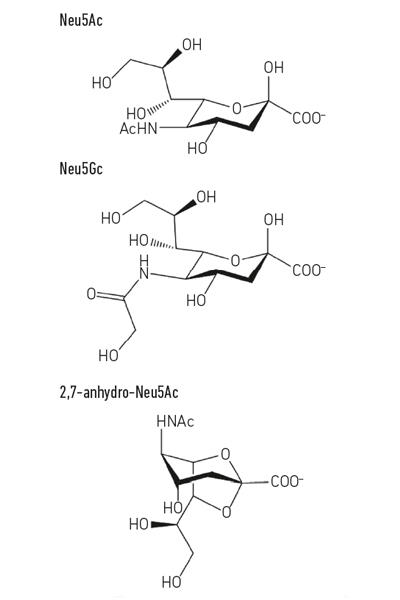
den vanligste sialinsyren, n-acetyl-neuraminsyre (Neu5Ac) (Fig. 1) er en relativt enkel sukkersyre som, bortsett fra ladningen, ikke har noe uvanlig uvanlig om det. Men når celler kontakter andre celler, bruker de ofte molekyler som vises på overflaten, og det er her sialinsyrer blir så viktige, da de vanligvis er de terminale sukkerene på vertsglykanene som pepperproteiner og lipider sitter på celleoverflater. Celleoverflate eller utskilt mucin glykoproteiner er gode eksempler på sialinsyrebelagte strukturer som mikrober kan støte på mukosale overflater, for eksempel luftveiene eller mage-tarmkanalen. Denne unike posisjonen som meet and greet-molekylet for andre humane celler, bakterieceller og virus setter sialinsyrene i sentrum for mange viktige prosesser.
- Fig. 1. Eksempel på sialinsyrederivater. Merk At n-glykolyl-neuraminic syre (Neu5Gc) ikke kan gjøres av mennesker, en biokjemisk forskjell som skiller oss fra andre store aper, men kan innlemmes i våre celleoverflater når de blir kjøpt fra dietten.
- Sukkerbelagte patogener
- Kryssmating-deling av søtsaker rundt
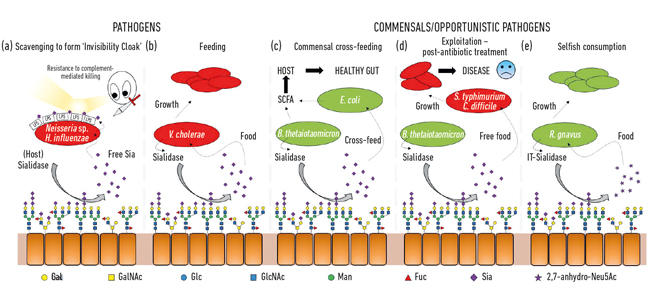
- Fig. 2. Tegneserie illustrasjon av de ulike interaksjonene mellom bakterier og sialinsyrer som er tilstede som terminale sukkerarter på en rekke vertsglykaner. Disse spaltes av sialidaser og konverteres vanligvis til fri Neu5Ac med unntak AV IT-sialidasen som frigjør 2,7-anhydro-Neu5Ac. Fri sialinsyre frigjort av verts-eller patogene sialidaser kan deretter brukes av patogener som en usynlighetskappe (a), eller til mating (b). Kryssmating kan forekomme mellom medlemmer av tarmmikrobiotaen eller mellom kommensale og patogene bakterier som deler samme nisje (c, d) mens andre bakterier har utviklet egoistiske mekanismer for å reservere sialinsyre til eget forbruk (e).
- Holde tak i dine egne søtsaker
- Virus kommer inn på loven
- Videre lesing
- Andrew Bell
- Emmanuele Severi
- Nathalie Juge
- Gavin H. Thomas
- hvorfor er mikrobiologi viktig?
- Hvilke råd vil du gi til noen som starter på dette feltet?
Fig. 1. Eksempel på sialinsyrederivater. Merk At n-glykolyl-neuraminic syre (Neu5Gc) ikke kan gjøres av mennesker, en biokjemisk forskjell som skiller oss fra andre store aper, men kan innlemmes i våre celleoverflater når de blir kjøpt fra dietten.
Sukkerbelagte patogener
En av de første mikrobielle funksjonene til sialinsyrer var forbundet med evnen til noen patogener å kolonisere og deretter forårsake sykdom hos mennesker. Faktisk pionerer ex-Presidenten For Mikrobiologiforeningen, Professor Harry Smith FRS, som jobber ved University Of Birmingham, dette arbeidet Med Neisseria gonorrhoea. Når forskere i hans gruppe isolerte feilen direkte fra kroppen, var den motstandsdyktig mot drap ved humant serum( komplementmediert drap), men hvis feilene ble dyrket i laboratoriet i noen generasjoner, ble denne beskyttelsen tapt og cellene ble drept av serumet. Hans gruppe hadde oppdaget at disse patogenene stjeler sialinsyre fra verten og bruker den til å belegge sin egen celleoverflate for å humanisere dem. Men da de bare kan scavenge denne sialinsyren, betyr fraværet i vekstmedier at det ikke tilsettes ytterligere sialinsyre, slik at de raskt mister denne serumbeskyttelsen og blir drept. Det viser seg at en rekke andre sneaky bugs bruker lignende strategier for å generere sin egen usynlighetskappe ved hjelp av sialinsyre, Som Neisseria meningitidis, Campylobacter jejuni eller Haemophilus influenzae(Fig . 2a)
Kryssmating-deling av søtsaker rundt
i tillegg til å bli brukt av patogener som en usynlighetskappe, er sialinsyrer ganske god mat for bakterier, da de gir både karbon og nitrogen, og kan brukes som energikilde. Tidligere arbeid viste at patogener som vibrio cholerae var glade for bare å spise sialinsyrer, og det var i seg selv viktig for vellykket vertskolonisering (Fig. 2b).
i tarmen kommer en viktig kilde til sialinsyrer fra muciner, som er de viktigste strukturelle komponentene i slimlaget som dekker epiteloverflaten. Det mest utsatte laget av slim er habitat for ulike kommensale bakterier, som har lært å feire på mucinproteiner, hvor sukker utgjør 80% av deres masse. Fra et enkelt topologisk perspektiv høstes sialinsyrene, som de terminale sukkerene, først, og utsetter de underliggende sukkerene som også senere kan frigjøres og spises. Med fremkomsten av neste generasjons sekvensering har vi fått kunnskap om mikrober som lever i denne nisje, og mange har gener for sialinsyreopptak og påfølgende katabolisme. Det som var spennende var oppdagelsen av at noen av de commensale anaerobene, som Bacteroides thetaiotaomicron, utskiller sialidaser for å frigjøre sialinsyren, men mangler transportører og katabolske gener for å faktisk bruke den. Denne tilsynelatende altruistiske bevegelsen er nødvendig for at bakteriene skal få tilgang til underliggende sukkerarter og underbygger en prosess med metabolsk kryssmating til andre bugs som lever i slimnissen, som commensal Escherichia coli-stammer som ikke har en sialidase, men er gode til å spise fri sialinsyre (Fig. 2c). Dette konseptet er nå anerkjent som et viktig aspekt ved kolonisering og nisjetilpasning. En strålende eksemplifisering av Dette ble beskrevet Av Gruppen Justin Sonnenburg fra Stanford University, ved hjelp av en musemodell med en enkel definert tarmmikrobiota. Etter antibiotikabehandling ble mange av de sialinsyrespisende bakteriene utryddet, noe som resulterte i akkumulering av fri sialinsyre i tarmen. Når de da utfordres med patogener Som Salmonella typhimurium eller Clostridium difficile, vil disse patogenene dra nytte av den frie sialinsyren som næringsstoff(Fig. 2d). Mutantstammer av patogener som mangler transportørene eller katabolske gener, hadde ingen fordel, noe som førte til en direkte forbindelse til patogen utvekst når det mikrobielle samfunnet i tarmen forstyrres. Mekanismer for å redusere bassenget av fri sialinsyre i tarmen ble derfor foreslått som en del av en ny antiinfeksjonsstrategi.
Fig. 2. Tegneserie illustrasjon av de ulike interaksjonene mellom bakterier og sialinsyrer som er tilstede som terminale sukkerarter på en rekke vertsglykaner. Disse spaltes av sialidaser og konverteres vanligvis til fri Neu5Ac med unntak AV IT-sialidasen som frigjør 2,7-anhydro-Neu5Ac. Fri sialinsyre frigjort av verts-eller patogene sialidaser kan deretter brukes av patogener som en usynlighetskappe (a), eller til mating (b). Kryssmating kan forekomme mellom medlemmer av tarmmikrobiotaen eller mellom kommensale og patogene bakterier som deler samme nisje (c, d) mens andre bakterier har utviklet egoistiske mekanismer for å reservere sialinsyre til eget forbruk (e).
Holde tak i dine egne søtsaker
mens mye tidlig sialomicrobiology fokusert på patogener, har nyere arbeid fokusert på commensal bakterier bebor samme nisje. Dagens studier innen tarmmikrobiota avslører hvor hard konkurransen Om Neu5Ac er i tarmen vår og hvordan den kan tippe balansen mellom patogener og kommensaler. Hva om en bestemt bakterie kunne frigjøre sialinsyre i en form som bare den kunne få tilgang til? Arbeid I Juge-Gruppen i Norwich viste at tarmsymbiont ruminococcus gnavus har utviklet seg til å gjøre det, ved hjelp av en spesiell type sialidase kalt en intramolekylær trans-sialidase (IT-sialidase)som frigjør 2,7-anhydro-Neu5Ac (Se Fig. 1) i stedet For Neu5Ac. Ved å utvikle en metode for å syntetisere ren 2,7-anhydro-Neu5Ac viste de At r. gnavus-stammer kunne vokse på 2,7-anhydro-Neu5Ac som eneste karbonkilde. I tillegg til domenet som katalyserer den enzymatiske reaksjonen, omfatter IT-sialidaser et karbohydratbindende domene som bidrar til å formidle bindingen AV IT-sialidasen til sialinsyrerike muciner. In vivo kan dette favorisere en mekanisme som bidrar til å målrette bakteriene mot sialinsyrerike regioner I GI-kanalen, og dermed fremme bakteriell kolonisering i slimlaget. Når den er plassert på rett sted, spalter IT-sialidasen av sialinsyre fra terminalkjedene av muciner og frigjør den i en form, 2,7-anhydro-Neu5Ac, som R. gnavus kan bruke Som sin egen matkilde, i stedet for å dele Neu5Ac Med andre bakterier som lever i slimnissen, inkludert patogener(Fig . 2e). Denne mekanismen gir en konkurransefortrinn over andre sialinsyrespisende bakterier som bor i slimnissen, ved å gi en kilde til næringsstoffer som den fortrinnsvis kan få tilgang til. I Tillegg kan r. gnavus også bidra til å redusere utveksten av enteriske patogener ved å redusere Nivået Av Neu5Ac i slimhinnemiljøet, og åpne måter til en ny terapeutisk strategi i en alder av økende antibiotikaresistens.
Virus kommer inn på loven
Virus bruker også sialinsyrer som nøkkelmolekyler under infeksjonssyklusen, mest kjent influensavirus. Initial vedlegg er det første nøkkeltrinnet da sialinsyrer fungerer som reseptorer for viruset på respiratoriske epitelceller. Typen av kobling av sialinsyren til underliggende sukker i glykanene styrer ofte vertsområdet for influensavirus, og pandemiske stammer har ofte adhesjonsprotein (hemagglutinin) som gjenkjenner de menneskelignende former for sialinsyrefeste og så er spesielle virulente. Sialinsyre er viktig i det siste stadiet i virussyklusen, da frigjøring fra den infiserte cellen krever spaltning av sialinsyrer fra vertscellens overflate. Dette katalyseres av et virusenzym som kalles sialidase (eller neuraminidase). Hemming av dette enzymet ved strukturelle analoger av sialinsyre, formulert I Tamiflu® (Oseltamivirfosfat) Og Zanamivir, er vår førstelinjebehandling for influensa og er en triumf av struktur/funksjon inspirert legemiddeldesign.som konklusjon er det klart at sialinsyrer er kritiske molekyler i mange prosesser hvor mikrobielle celler og virus interagerer med pattedyrcelleoverflater. I denne korte artikkelen har vi berørt noen av disse for å gi en ide om de sentrale funksjonene til disse sukkene i kommunikasjon, stealth, kolonisering og metabolisme i ulike aspekter av hede og sykdom. Sialinsyrer har blitt beskrevet som de viktigste kjemikaliene i verden og vil uten tvil fortsette å avsløre mer om utviklingen av mennesker og deres samspill med deres mikrober.
Videre lesing
Thomas, GH. Sialinsyreoppkjøp i bakterier – ett substrat, mange transportører. Biochem Soc Trans 2016; 44: 760-765.Ng KM, Ferreyra JA, Higginbottom SK, Lynch JB, Kashyap PC et al. Mikrobiota-frigjorte vertssukker letter post-antibiotisk utvidelse av enteriske patogener. Natur 2013; 502: 96-99.
Owen CD, Tailford LE, Monaco S, Hryvuligoj T, Vaux L et al. Unraveling spesifisitet og mekanismen av sialinsyre anerkjennelse av tarmen symbiont Ruminococcus gnavus. Nat Commun 2017; 8: 2196.Tailford LE, Owen CD, Walshaw J, Crost EH, Hardy-Goddard J, Le Gall G, De Vos WM, Taylor GL, Juge N. Oppdagelsen av intramolekylære trans-sialidaser i menneskelig tarmmikrobiota antyder nye mekanismer for mukosal tilpasning. Nat Commun 2015; 6: 7624.
Juge N, Tailford L, Owen CD. Sialidaser fra tarmbakterier: en mini-gjennomgang. Biochem Soc Trans 2016; 44:166-175.

Andrew Bell
Quadram Institute Bioscience, Norwich Research Park, NR4 7UQ, UK
Andrew Bell er en post graduate student I Avdelingen Gut Mikrober og Helse Ved Quadram Institute studere virkningen av en ny mekanisme for sialinsyre metabolisme på gut homeostase. Han fikk Sin BSc I Biokjemi fra University Of East Anglia og tilbrakte 2 år på John Innes Centre på klorofyllmetabolisme i pea (Pisum sativum) før Han ble Med I Nathalie Juges gruppe Ved Quadram Institute i 2015 for sitt doktorgradsarbeid.

Emmanuele Severi
Institutt For Biologi, University Of York, Wentworth Way, York YO10 5DD, STORBRITANNIA
@emm_severi
Emmanuele Severi er en postdoktor ved Institutt For Biologi ved University Of York som arbeider i laboratoriet Til Prof Gavin Thomas. Han har jobbet med sialinsyretransport og metabolisme på ulike tidspunkter i sin karriere, og har vært Medlem av Samfunnet siden 2002.

Nathalie Juge
Quadram Institute Bioscience, Norwich Research Park, NR4 7UQ, UK
Nathalie Juge Er Forskningsleder ved Quadram Institute Bioscience (QIB) I Norwich, Nestleder For Instituttets Strategiske Program, Gut Microbes and Health (GMH), Og Æresprofessor Ved School Of Biological Sciences, University Of East Anglia. Hun leder En Forskningsgruppe på glykobiologi av vert-mikrobe-interaksjoner i tarmen med fokus på mucin-avledet sialinsyremetabolisme av tarmmikrobiotaen (https://quadram.ac.uk/nathalie-juge).

Gavin H. Thomas
Institutt For Biologi, University Of York, Wentworth Way, York YO10 5DD, STORBRITANNIA
@GavinHThomas
Gavin H. Thomas Er Professor I Mikrobiologi Ved Institutt For Biologi Ved University Of York og har jobbet med bakteriell sialinsyretransport og katabolisme for over et tiår. Han har vært Medlem av Foreningen siden 1995, Var Redaktør For Microbiology Today (2003-2006), tjenestegjorde I Rådet i samme periode, tjenestegjorde I Communications committee (2012-2015) og Equality & Diversity committee (2013-2015). Han har Vært Redaktør, Seniorredaktør og Er Nå Assisterende Redaktør I Samfunnets ledende tidsskrift Microbiology (http://thomaslabyork.weebly.com).
hvorfor er mikrobiologi viktig?
Gavin: Det er viktig for helsen vår – først nå begynner vi virkelig å forstå den molekylære sammensetningen og funksjonen til mikrobiomet vårt, noe som sannsynligvis påvirker kroppen vår på mange måter.
Hvilke råd vil du gi til noen som starter på dette feltet?
Gavin: Les mye og utover ‘din feil’ – mikrober kan gjøre stort sett alt og har vanligvis oppfunnet flere forskjellige måter å gjøre det på!
Bilder: H3N2 influensavirus partikler. CDC / Science Foto Bibliotek.
Fig. 1. Andrew Bell.
Fig. 2. Emmanuele Severi.