Pyruvatkinase m2 brensel flere aspekter av kreftceller: fra cellulær metabolisme, transkripsjonell regulering til ekstracellulær signalering
Biokjemisk analyse ved å karakterisere den enzymatiske aktiviteten som katalyserer dannelsen av laktat fra glukose i cellelysater avslørte den første intracellulære metabolske banen, glykolytisk vei. Fra rensingen av fraksjoner som inneholdt glykolytisk aktivitet, bidro en rekke pionerforskere til identifisering av enzymer som involverer i hvert trinn i banen . Disse resultatene bygger opp vårt moderne konsept i utveksling av aerob og anaerob respirasjon og energiproduksjon under ulike fysiologiske og patologiske forhold.eksistensen av et enzym som katalyserte PRODUKSJONEN AV ATP ved å overføre en fosfatgruppe fra PEP TIL ADP i leveren ble først rapportert i 1934 . Etterfølgende isolering av enzymet, kjent som pyruvatkinase (PK) senere, viste forskjeller i vevsfordeling og katalytisk kinetikk som tyder på at dette enzymet kan ha forskjellige isoformer . I løpet av 1982 til 1984 ble ULIKE PK-gener klonet fra gjær, kylling og rotte . Den funksjonelle studien AV PKM2 ble initiert ved identifisering av et kandidatgen i mus tidlig på 1980-tallet . Senere, Noguchi et al. viste at to isoformer AV PK (PKM1 og PKM2) er kodet av det samme pkm-genet via alternativ spleising . Hos mennesker produseres OGSÅ PKM-isoformer via en lignende spleisingsmekanisme ved å inkludere ekson 9 og 10 i PKM1 og pkm2 mRNA separat .
Flere funn fanget forskerens oppmerksomhet på PKM2S potensielle rolle i tumorigenese. FOR DET første ER PKM2 den embryonale isoformen som uttrykkes sterkt under dyrs utvikling. Transkripsjonen dempes i en rekke voksne vev mens den reaktiveres i svulster . For det andre viste studier av den relative overfloden AV PKM1 og PKM2 i normalt vev og tumorvev en overgang fra pkm1-isoformen til PKM2-isoformen ved ulike kreftformer som hepatocellulært karsinom . For det tredje forsterkes endringen av mRNA-spleising FRA PKM1 TIL PKM2 av c-Myc onkogen som tyder på at kreftceller aktivt engasjerer seg i denne bryteren for å passe deres krav i proliferasjon og metabolisme . For det fjerde påvirker modulering AV PKM2-aktivitet av aktivatorer eller hemmere tumorvekst in vivo .
- den første episoden: PKM2 som et metabolsk enzym i cytoplasma
- den andre episoden: pkm2 som en signalmodulator i cytoplasma
- den tredje episoden: PKM2 som en transkripsjonell regulator i kjernen
- den fjerde episoden: PKM2 som ekstracellulær signalkommunikator
- The fifth episode: ubesvart avvik AV PKM2
- den pågående episoden: ER PKM2 en kreftbiomarkør og narkotikamål?
den første episoden: PKM2 som et metabolsk enzym i cytoplasma
Siden PKM2S rolle i metabolsk kontroll av glykolyse i kreftceller har blitt grundig gjennomgått, oppsummerer vi bare tre viktige forskjeller mellom PKM1 – og PKM2-mediert katalyse og cellulær metabolisme her. Den første forskjellen er underenhet interaksjon. BÅDE PKM1 OG PKM2 er tetrameriske proteiner dannet av fire identiske underenheter. Hver underenhet (eller monomer) inneholder fire strukturelle domener, inkludert a, B, C og N-terminal domene. Monomeren først dimerizes sammen og deretter to dimerer samhandle via dimer-dimer grensesnitt orkestrert av c-domenet til monomer for å danne en tetramer. FORDI PKM1 og PKM2 inkluderer forskjellige eksoner i deres mrna, endrer dette de kodede aminosyrene i C-domenet og endrer tetramerstabiliteten. Under fysiologisk tilstand organiserer PKM1 konstitusjonelt som en tetramer mens PKM2 kan eksistere i tetramer eller dimer. Den andre forskjellen er allosterisk regulering. Avhengig av intracellulære konsentrasjoner av små molekyler og metabolitter, kan AKTIVITETEN TIL PKM1 og PKM2 reguleres differensielt. En av de mest kjente allosteriske regulatorene er fruktose-1,6-bisfosfat (FBP). Dette glykolytiske mellomproduktet binder PKM2 direkte og øker affiniteten TIL PKM2 for PEP . TVERT imot påvirker FBP ikke signifikant pkm1-aktivitet. I TILLEGG TIL FBP har andre metabolitter, aminosyrer og små molekyler blitt rapportert å påvirke PKM2-aktivitet (Fig. 1). Konsentrasjonen som kreves for aktivering eller hemming er imidlertid høy og den modulerende effekten er beskjeden. Hvorvidt disse små molekylene spiller en viktig rolle i kontrollen AV PKM-aktivitet under fysiologiske forhold, er fortsatt uklart. PKM2-aktivitet reguleres også ved post-translasjonell modifikasjon, som fosforylering, acetylering og oksidasjon, som favoriserer den lave aktiviteten til dimerisk PKM2(Fig. 1). Den tredje forskjellen er energiproduksjon og mellomutnyttelse. SIDEN PKM1 konstitutivt eksisterer som den aktive tetrameren, er DEN viktigste biologiske funksjonen TIL denne isoformen genereringen AV ATP for å levere cellulær energi. PKM2, i tillegg til å produsere ATP, kan imidlertid bytte til den mindre aktive dimeriske formen for å generere flere glykolytiske mellomprodukter som kan brukes som byggesteiner for biosyntese av aminosyrer, lipider og nukleotider.
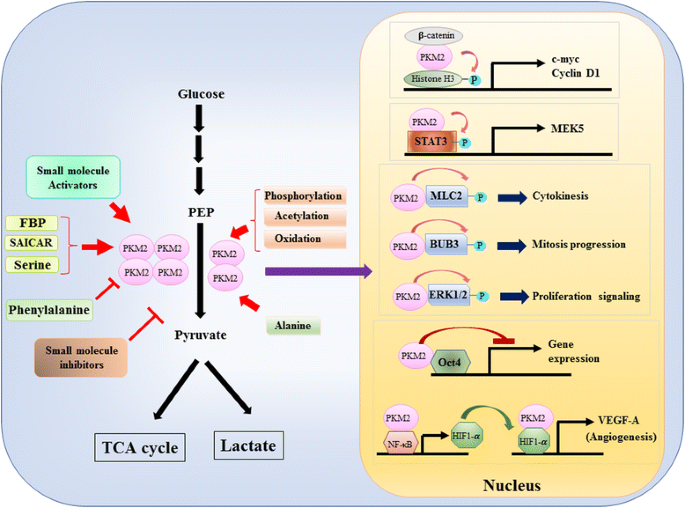
Modulering AV PKM2 aktivitet ved fysiologiske molekyler og post-translasjonell modifikasjon og DEN potensielle rollen TIL PKM2 i transkripsjonell regulering
den andre episoden: pkm2 som en signalmodulator i cytoplasma
i tillegg til å fungere som et glykolytisk enzym, foreslås pkm2 å involvere i flere cellulære prosesser på Grunn av Identifisering av INTERAGERENDE PROTEINER i cytoplasma. FOR eksempel ble PKM2 vist å være et interagerende protein av flere tyrosinkinaser, inkludert A-Raf, Break point cluster region-Abelson (BCR-ABL) fusjonskinase, fibroblastvekstfaktorreseptor 1 (FGFR1) etc. . Disse bindingspartnerne har vist seg å modulere den dimere / tetrameriske endringen AV PKM2 for å endre cellemetabolismen. DET er imidlertid mulig AT PKM2 gjensidig kan påvirke katalytisk kinetikk, substratbinding og cytoplasmatisk plassering av disse bindingspartnerne for å modulere signaltransduksjon. Funnet at PKM2 er et fosfor-tyrosinbindende protein styrker denne muligheten fordi mange intracellulære signalmediatorer kan binde seg til fosfortyrosinrester for å samle spesifikke proteinkomplekser for signaloverføring . Hittil vokser listen over pkm2 bindende partnere kontinuerlig. Vi fremhever flere nye medlemmer og diskuterer deres biologiske implikasjon her. Mukheriee et al. viste AT PKM2 kunne binde Seg Med HuR, ET rna-bindende protein som spiller en viktig rolle i kontrollen av mrna-stabilitet og translasjonseffektivitet, for å fremme cellesyklusprogresjon og proliferasjon av gliomceller . Interessant nok ble et ANNET rna-bindende protein tristetraprolin som kunne binde et antall mRNA via DET AU-rike elementet i 3′-utranslasjonal region (3′ – UTR) også funnet å være EN PKM2-interaksjonspartner, OG PKM2 induserte fosforylering og nedbrytning av tristetraprolin for å modulere brystkreftvekst . Disse to studiene antyder en potensiell translasjonell kontrollfunksjon AV PKM2. Nylig, Liang et al. identifisert det anti-apoptotiske proteinet Bcl2 som en ny pkm2-partner . De viste at oksidativt stress induserte translokasjonen AV PKM2 til mitokondrier hvor den fosforylerte Og stabiliserte Bcl2 ved å forhindre nedbrytning via ubiquitineringsavhengig vei. Disse dataene antydet AT PKM2 hjelper kreftceller til å tilpasse oksidativt stress fremkalt av intracellulær metabolsk forandring eller ekstracellulær fornærmelse.
den tredje episoden: PKM2 som en transkripsjonell regulator i kjernen
en nukleær rolle AV PKM2 i reguleringen av gentranskripsjon eller epigenetisk modifikasjon ble først foreslått ved funnet AT PKM2 bundet Med Y333-fosforlert β-catenin, og β-catenin-pkm2-komplekset ble rekruttert til nukleosomene for å fosforylere histon H3 ved treonin 11 (Fig . 1). Denne fosforyleringen økte deretter histon H3-acetylering som førte til oppregulering av β-catenin målgener. En annen transkripsjonsfaktor direkte fosforylert AV PKM2 er signaltransduser og aktivator av transkripsjon 3 (STAT3). PKM2-mediert fosforylering AV STAT3 ved tyrosin 705 forbedret STAT3-aktivitet for å oppregulere ekspresjonen av mitogenaktivert proteinkinasekinase 5 (MEK5). I tillegg til transkripsjonsfaktorer er PKM2 vist å fosforylere myosin light chain 2 (MLC2), BUB3 og ekstracellulær signalregulert kinase 1 og 2 (ERK1 og ERK2). Interessant, PKM2 virker også via fosforyleringsuavhengig måte å påvirke genuttrykk. FOR eksempel har PKM2 blitt funnet å binde Med Oct4, en av de viktigste transkripsjonsfaktorene som styrer selvfornyelse av stamceller, og hemmer Oct4-mediert transkripsjon . PKM2 kan også øke tumorangiogenesen ved å interagere MED NF-kB og HIF-1Α i kjernen og aktivere uttrykket FOR HIF-1Α målgen VEGF-A. økt sekresjon av VEGF-A øker følgelig dannelsen av blodkar som bidrar til tumorvekst . Selv om disse studiene sterkt antydet nukleær lokalisering og proteinkinase funksjon AV PKM2 i ulike fysiologiske og patologiske forhold, men betydningen av nukleær PKM2-mediert genuttrykk har blitt utfordret av studier ved hjelp AV pkm2 knockout celler. Ved å bruke-merket PEP og PKM2-null mus embryonale fibroblaster, Hosios et al. viste AT PEP-avhengig fosforylering ikke er en vanlig hendelse i celler, og reaksjonen katalyseres ikke AV PKM2 . Uoverensstemmelsen i disse studiene er for tiden uløst, og PROTEINKINASEAKTIVITETEN til PKM2 trenger ytterligere bekreftelse.
den fjerde episoden: PKM2 som ekstracellulær signalkommunikator
tilstedeværelsen av ekstracellulær PKM2 åpner en ny avenue for studiet AV pkm2 biologisk funksjon. Buschow et al. gitt det første beviset på AT PKM2 kunne påvises i b-celleeksosomer og ble identifisert som ET MHC klasse II-assosiert protein . To påfølgende studier indikerte også AT PKM2 eksisterte i eksosomer utgitt av forskjellige celler . For tiden gir flere offentlige databaser som ExoCarta og EVpedia omfattende informasjon for komponentene, inkludert proteiner, lipider, nukleinsyrer av ekstracellulære vesikler i forskjellige arter. Alle dataene bekrefter AT PKM2 er et pakkeprotein av eksosomer. Nylige studier har tydelig vist en kommunikativ rolle av eksosomer ved å levere forskjellige komponenter fra vertsceller til mottakerceller . DET er forventet AT PKM2 kan spille en rolle i celle-celle crosstalk.
Fremvoksende bevis støtter faktisk denne hypotesen. For eksempel viste en nylig studie at blod sirkulerende PKM2 kan fremme tumorvekst og angiogenese ved å øke veksten, migrasjonen og matriksadhesjonen av endotelceller . En annen undersøkelse viste også AT PKM2 utskilt fra kolonkreftceller kan virke via en autokrin stimulering for å øke cellemigrasjonen ved å aktivere pi3k/Akt-og wnt/β-catenin-veiene . I tillegg til kreftceller kan nøytrofiler på vevsskadestedene frigjøre PKM2 for å fremme angiogenese og sårheling . Vår siste studie viste også at rekombinant pkm2-protein kunne indusere fosforylering og aktivering AV epidermal vekstfaktorreseptor (EGFR). Videre fant VI AT r339e mutant PKM2 som fortrinnsvis dannet dimerisk PKM2 aktiverte EGFR mer signifikant enn den tetrameriske PKM2. Keller et al. identifiserte 154 proteiner som potensielle substrater FOR PKM2 etter behandling Av Hela-celler med succinyl-5-aminoimidazol-4-karboksamid-1-ribose-5′ – fosfat (SAICAR), en intracellulær metabolitt som kan stimulere PROTEINKINASEAKTIVITETEN TIL PKM2 . DE fant OGSÅ EGFR som et pkm2-substrat. Deres resultater er forskjellige fra våre på to måter. For det første utløses signalveiene som er aktivert i studien av ekstracellulær PKM2, mens de molekylære målene som er identifisert i studien, er potensielle substrater av intracellulær PKM2. For det andre initieres økning AV ERK1/2-aktivitet i vår studie VED egfr-aktivering, MENS ERK1 / 2-aktivering i studien stimuleres direkte av saicar / PKM2-komplekset. Et lignende fenomen observert i begge studiene er AT r339e mutant PKM2 aktiverer signalmolekyler mer signifikant enn den VILLE TYPEN PKM2 som tyder på den distinkte rollen dimeric OG tetramer PKM2 har i onkogenesen. Ved å bruke reseptortyrosinkinasearray fant vi at ekstracellulær PKM2 bare aktiverte begrensede vekstfaktorreseptorer i brystkreftceller (data ikke vist). For tiden er selektiviteten av reseptoraktivering av ekstracellulær PKM2 ukjent. I tillegg er hvorfor r339e mutant PKM2 er mer potent i aktiveringen AV EGFR heller ikke klart. Flere eksperimenter er nødvendig for å svare på disse spørsmålene.Et annet elegant spørsmål som skal tas opp er om fri pkm2 OG vesikelemballasje PKM2 har lignende effekt for å fremme tumorigenese (Fig. 2). Fritt ekstracellulært PKM2 kunne ikke trenge gjennom plasmamembranen og kunne bare aktivere intracellulær signalering via celleoverflateproteiner som vekstfaktorreseptorer. Omvendt kan vesikelemballasje PKM2 være endocytosed av kreft og stroma celler, OG PKM2 frigjort fra vesikler kan påvirke metabolisme og genuttrykk via intracellulær mekanisme. The PKM2-null cells or mice will be useful to elucidate whether these two extracellular forms of PKM2 could act synergistically or antagonistically in tumorigenesis.

The potential tumor-promoting effect of extracellular PKM2
The fifth episode: ubesvart avvik AV PKM2
i tillegg til de cellebaserte dataene som er omtalt ovenfor, har den onkogene rollen TIL PKM2 også blitt utfordret etter genereringen AV PKM2 knockout-mus. Israelsen et al. generert en betinget knockout musemodell ved å slette PKM2-spesifikke exon 10 . Overraskende nok akselererte uttømming AV PKM2, men ikke svekket tumordannelse drevet av Tap Av Brca1-gen hos mus. Disse dataene indikerte AT PKM2 ikke er nødvendig for spredning av kreftceller. Interessant NOK ble pkm1-uttrykk bare påvist i ikke-prolifererende tumorceller som tyder på en tumor-undertrykkende rolle AV PKM1 i brystkreft. I TILLEGG har PKM2 knockout mus en høy forekomst for å utvikle hepatocellulært karsinom spontant etter lang ventetid på grunn av ubalansen i metabolisme . Disse resultatene mot forestillingen OM AT PKM2 spiller en onkogen rolle in vivo.
den pågående episoden: ER PKM2 en kreftbiomarkør og narkotikamål?
selv om resultatene av genmodifisert musemodell ikke støtter DEN tumorfremmende aktiviteten TIL PKM2, er overekspresjon av PKM2 universelt funnet i humane kreftformer og er assosiert med dårlig klinisk utfall (Tabell 1) . To nyere meta-analysestudier støttet også denne konklusjonen. Wu et al. analyserte data fra 2812 pasienter med solide svulster i fordøyelsessystemet oppnådd fra 16 kohortstudier og fant at overuttrykk AV PKM2 er assosiert med redusert total overlevelse i magekreft, esophageal squamouscellekarsinom, hepatocellulært karsinom, biliær kreft og oral kreft . PKM2 er imidlertid ikke en prognostisk faktor for kreft i bukspyttkjertelen. Dette funnet er ikke i samsvar med tidligere studier som viser at økning AV PKM2 er en indikator på dårlig overlevelse hos pasienter med kreft i bukspyttkjertelen . En annen undersøkelse inkluderte 4796 tilfeller fra 27 individuelle studier som viste AT pkm2 oppregulering er korrelert med dårligere totaloverlevelse, sykdomsfri overlevelse og residivfri overlevelse i samlede data . PKM2, stratifisert etter krefttype, forutsier imidlertid ikke en dårlig overlevelse av kreft i bukspyttkjertelen. SAMLET SETT ser PKM2 ut til å være en ansvarlig prognostisk markør i de fleste solide svulster.
tvert imot er bruken AV PKM2 som diagnostisk faktor kontroversiell. En proteomisk analyse viste at PKM2 er en potensiell diagnostisk markør for påvisning av lungekreft . En nylig studie foreslo IMIDLERTID AT PKM2 ikke er en god diagnostisk markør for lungekreft på grunn av lav spesifisitet . PÅ SAMME måte er PKM2 alene usannsynlig å være en nyttig markør for screening av tykktarmskreft . Kombinasjonen av flere markører kan imidlertid øke sensitiviteten og spesifisiteten for kreftdiagnose .
det terapeutiske potensialet TIL PKM2 er en spennende hendelse i kreftbehandling. På den ene siden forventes hemming AV PKM2 å hemme glykolyse, svekke gentranskripsjon og undertrykke celleproliferasjon. DERFOR SYNES PKM2-hemmere å være gode kandidater for utvikling av legemidler mot kreft. Ved å bruke bibliotek screening, Vander Heiden et al. identifiserte tre nye klasser AV pkm2-hemmere og viste at den mest effektive forbindelsen hemmet PKM2-aktivitet og induserte død av kreftceller . Nylig, Ning et al. funnet at nye naftokinonderivater er potente pkm2-hemmere . En effektiv forbindelse 3 k undertrykte spredning av flere kreftcellelinjer ved sub-mikromolære konsentrasjoner mens den viste liten skadelig effekt på normale celler. Fra den andre siden kan aktivering AV PKM2 også hemme tumorvekst. FORDI DEN lave aktiviteten PKM2 dimer er den viktigste isoformen som utløser glykolyse i cytoplasma og gentranskripsjon i kjernen i kreftceller, KAN PKM2 aktivatorer som kan fremme dannelsen av tetramer PKM2 bytte glykolyse til mitokondrier vei og redusere kjernefysisk oppføring for å dempe gentranskripsjon. Begge effektene svekker metabolsk etterspørsel og vekststøttende signalering som fører til tumorregresjon. To pionerstudier identifiserte ulike pkm2-aktivatorer og karakteriserte deres spesifisitet in vitro . En senere studie viste at pkm2-aktivatorer faktisk fremmet tetramerdannelse og undertrykt tumorvekst in vivo . Disse resultatene antydet AT pkm2-aktivatorer kunne være lovende anti-kreftmedisiner.
Motstand mot kjemoterapi er en stor blokkering for kreftbehandling. Å overvinne den iboende kjemoresistansen av kreftceller er presserende for klinisk forskning. ROLLENE TIL PKM2 i kjemoresistens av kreftceller har blitt avslørt, og målretting AV PKM2 har vist seg å re-sensibilisere kjemoresistente kreftceller. EN nylig studie viste AT CD44 interagerer MED PKM2 og undertrykker PKM2-aktivitet via økende tyr105-fosforylering AV PKM2. CD44-ablasjon induserte overgangen fra aerob glykolyse til mitokondriell respirasjon og økende produksjon av reaktive oksygenarter (ROS), noe som resulterte i forbedring av cisplatinfølsomhet i kolorektale kreftceller . Hemming AV PKM2-aktivitet ble vist å undertrykke glykolyse og overvinne cisplatinresistens. Kombinasjonen av cisplatin og PKM2-hemmere kan derfor være en effektiv strategi for kjemoterapi (Fig. 3). PKM2 har også blitt rapportert å delta i reguleringen av gemcitabinresistens i pankreaskreftceller. Kim et al. viste AT PKM2 fremmer gemcitabinresistens gjennom negativ regulering av p38-mediert p53 fosforylering, og reduserer derfor transkripsjonell aktivitet av p53 og undertrykker ekspresjonen av pro-apoptotiske gener . Videre øker ektopisk ekspresjon AV R399E-PKM2, som fortrinnsvis danner dimerisk PKM2, motstanden av kreft i bukspyttkjertelen til gemcitabin. Disse bevisene antyder AT PKM2 bidrar til induksjon av legemiddelresistens via en ikke-metabolsk mekanisme. Fordi dimerisk PKM2 hovedsakelig virker som en transkripsjonell regulator i kreftceller, KAN PKM2-aktivatorer som fremmer dannelsen av tetramerisk PKM2 være effektive for å undertrykke ikke-metabolsk funksjon AV PKM2 og redusere resistens mot gemcitabin i pankreas kreftceller. Nuclear PKM2 har også vist seg å bidra TIL RESISTENS AV EGFR-hemmer i kolorektal kreft og lungekreft . Li et al. viste at nukleær PKM2-mediert STAT3 fosforylering reduserer følsomheten av kolorektale kreftceller til gefitinib og forstyrrelse av samspillet mellom nukleær PKM2 OG STAT3 gjenopprettet gefitinib følsomhet i cellene . Mer nylig viste en annen studie AT PKM2 translokerer inn i kjernen og interagerer med poly-ADP ribose (PAR) ved vekstfaktorstimulering. PARBINDINGSAKTIVITETEN TIL PKM2 er kritisk for nukleær retensjon og gentranskripsjon AV PKM2 og er viktig for å fremme celleproliferasjon og tumorvekst. De konkluderte med at inhibering AV pkm2 kjernefunksjon kan overvinne motstanden TIL EGFR-muterte kreftceller . Begge studiene pekte på en avgjørende rolle for kjernefysisk PKM2 i å formidle stoffresistens i kreft og foreslo at målretting av kjernefysisk PKM2 kan være en lovende strategi for å overstyre motstanden.

PKM2 bidrar til spredning og legemiddelresistens i kreftceller