células Gliais da Retina por Helga Kolb
Helga Kolb
Três tipos básicos de célula glial são encontradas na retina humana, células de Muller, astroglia e microglia. Todos foram descritos para a retina por Cajal há mais de cem anos (1892).1. Células Muller.as células Muller são a principal célula glial da retina. Eles formam estruturas de suporte arquitetônico que se estendem radialmente através da espessura da retina e são os limites da retina na membrana limitante externa e interna, respectivamente. Um entendimento completo da forma de uma célula Muller é melhor visto após a coloração de Golgi como mostrado originalmente por Cajal (1892) abaixo.
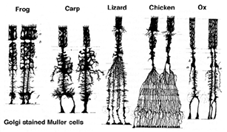
Fig. 1. As células Muller manchadas de Golgi situam-se na camada nuclear interna e projectam processos irregulares, espessos e finos em qualquer direcção até à membrana limitante exterior e à membrana limitante interna. Os processos celulares de Muller se insinuam entre os corpos celulares dos neurônios nas camadas nucleares e os grupos de envelopes dos processos neurais nas camadas plexiformes (Fig. 1). Na verdade, os processos neurais retinianos só são permitidos contato direto, sem envolver processos de células Muller, em suas sinapses.uma única célula progenitora dá origem tanto às células Muller como aos neurónios da retina (Turner e Cepko, 1987), embora aparentemente em duas fases. Os neurônios de fase mais antiga nascidos na margem apical do neuroepitélio adjacente ao pigmento epitélio produzem neurônios primários consistindo de cones, células horizontais e células ganglionares (Fig. 2, direita). A segunda fase das células também nascidas nas margens apicais produz células Muller e fotorreceptores rod, células bipolares e células de amacrina (Reichenbach e Robinson, 1995) (Fig. 2, left). Todos os neurônios em desenvolvimento e as células Muller têm que migrar para dentro de sua posição final e acredita-se que os processos e troncos de células Muller guiam grande parte das migrações de neurônios e dirigem as diferenciações neuróticas.
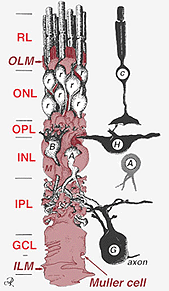
Fig. 2. Desenho esquemático 3-D da relação entre a célula Muller e outros neurónios retinianos
As junções que formam a membrana limitante exterior estão entre as células Muller e outras células Muller e as células fotorreceptoras como desmossomas resistentes ou adesivos da zonula. Em algumas espécies junções gap (especializada membrana associações e canais que permitem a passagem de pequenas moléculas e íons) ou junções apertadas são parte desses Muller junções celulares (Miller e Dowling, 1970), mas não tão em espécies de mamíferos, onde nenhuma tintura de acoplamento já foi observado (Robinson et al., 1993; Reichenbach and Robinson, 1995). A superfície da célula Muller face ao pigmento epitélio e espaço subretinal é expandida por muitas projeções da membrana celular Muller conhecida como vilosidades apicais. A membrana limitante interna, por outro lado, é formada pelo endfeet cónico da célula de Muller, mas não se vêem aqui junções especializadas. As células Muller também formam endfeet nos grandes vasos sanguíneos da retina na superfície interna da retina. A superfície da membrana celular Muller virada para o vítreo é coberta com um material mucopolissacarídeo e, portanto, forma uma verdadeira membrana basilar.as células Muller contêm glicogénio, mitocôndria e filamentos intermédios imunoreativos para a vimentina e, em certa medida, para a acidicproteína fibrilar glial (GFAP). Estes últimos filamentos encontram-se normalmente na metade interna das células Muller da retina e no seu endfeet (Fig. 3, esquerda), mas após trauma na retina, como descolamento da retina, tanto a vimentina e GFAP são massivamente re-regulados e Encontrados em toda a célula(Fig. 3, right) (Guerin et al., 1990; Fisher e Lewis, 1995).
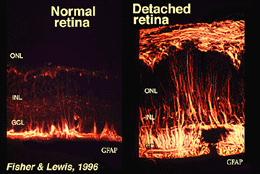
Fig. 3. Imunoreactividade GFAP em células Muller
as células Muller têm uma gama de funções, todas elas vitais para a saúde dos neurónios retinianos. As células Muller funcionam em uma relação simbiótica com os neurônios (para uma excelente revisão veja Reichenbach e Robinson, 1995). Assim, as funções das células Muller incluem:
- fornecendo produtos finais do metabolismo anaeróbico (degradação do glicogénio) para alimentar o metabolismo aeróbico nas células nervosas.limpam os resíduos neurais, como o dióxido de carbono e o amoníaco, e reciclam os transmissores de aminoácidos usados.protegem os neurónios da exposição a neurotransmissores em excesso, como o glutamato, utilizando mecanismos de absorção bem desenvolvidos para reciclar este transmissor. Eles são particularmente caracterizados pela presença de altas concentrações de glutamina sintase.podem estar envolvidos tanto na fagocitose dos detritos neuronais como na libertação de substâncias neuroactivas tais como GABA, taurina e dopamina.
- Eles são pensados para sintetizar retinoic de retinol (retinoic é conhecido por ser importante no desenvolvimento do olho e o sistema nervoso) (Edwards, 1994)
- Que controlam a homeostase e proteger os neurônios da deletérios alterações em suas iônica ambiente, tomando-se extracelular de K+ e redistribuindo-a.
- Que contribuem para a geração do electroretinogram (ERG) b-onda (Miller e Dowling, 1970; Newman e Odette, 1984), a lenta P3 componente do ERG (Karwoski e Proenza, 1977) e o escotópica limiar de resposta (STR) (Frishman e Steinberg, 1989). Eles fazem isso através da regulação da distribuição K+ através da borda vítrea da retina, através de toda a retina e localmente na camada plexiforme interna da retina (Fig. 4, from Reichenbach and Robinson, 1995, adapted from Newman, 1989).

Fig. 4. Regulação de K+ por células Muller em células Muller
2. Astrocitos.
astrócitos não são células gliais do neuroepitélio da retina, mas entram na retina em desenvolvimento a partir do cérebro ao longo do nervo óptico em desenvolvimento (Stone e Dreher, 1987; Chan-Ling 1994). Eles têm uma morfologia característica de um corpo celular achatado e uma série fibrosa de processos de irradiação. Filamentos intermédios enchem os seus processos e, assim, mancham dramaticamente com anticorpos contra o GFAP (Schnitzer, 1988). Corpos e processos de células astrocíticas são quase inteiramente restritos à camada de fibra nervosa da retina. Sua morfologia muda da periferia para a cabeça do nervo óptico: a partir de uma forma de stellate simétrica na retina periférica (figos. 5a and b) (Schitzer, 1988) to extremely elongated near the optic nerve (Fig. 6 e 7).
 Fig. 5-a. astrócitos na retina periférica. Schnitzer, 1988 |
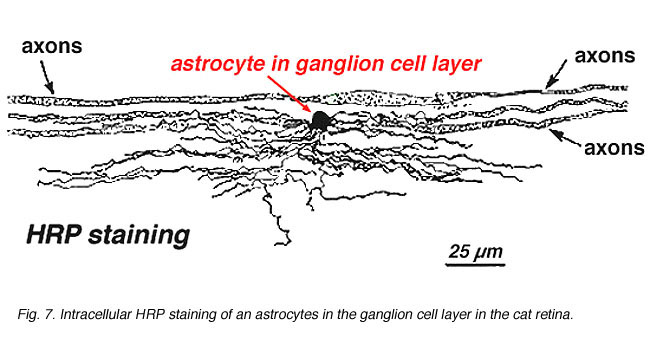
na coloração imunocitoquímica(Fig. 5. 5) e nas injecções intracelulares de HRP(Fig. 7) astrócitos manchados exibem claramente processos alinhados ao longo dos axônios de células ganglionares que percorrem a camada de fibra nervosa. Na distribuição, os astrócitos atingem o seu pico na cabeça do nervo óptico e têm um declínio relativamente uniforme na densidade em anéis radiantes da cabeça do nervo. Não estão presentes na avascular fovea ou ora serrata.
 Fig. 6. Astrocitos na retina central. Schnitzer, 1988 |
astrocitos espessos e finos foram distinguidos com base na coloração com anticorpos contra GFAP (Trevino et al., 1996). Assim, os astrócitos são dispostos sobre a superfície da célula de ganglion agrupam-se à medida que se dirigem para a cabeça do nervo óptico formando um tubo através do qual os axônios correm (Fig. 8). Junções de Gap e junções de adesão de zonula foram descritas entre processos astrocíticos em retina de gato (Höllander et al., 1991).
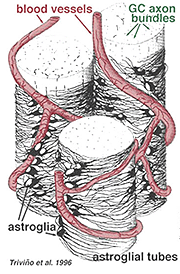
Fig. 8. O bloco 3-D de astrócitos dispostos sobre a superfície de gânglios de células axon agrupa
os vasos sanguíneos que circulam dentro e entre os gânglios de células também são cobertos por ambos os processos e mesmo por um corpo celular ocasional de um astrocito. A função dos astrócitos que envolvem os axônios de gânglios de células e a relação com os vasos sanguíneos da camada de fibra nervosa sugere que são bainhas axonais e vasculares gliais e parte de uma barreira hemato-encefálica. Similar às células Muller, elas são conhecidas por conter glicogênio abundante e podem formar um serviço nutritivo no fornecimento de glicose para os neurônios. Além disso, eles provavelmente servem um papel na homeostase iônica na regulação dos níveis de potássio extracelular e metabolismo de neurotransmissores como GABA.3. Células microgliais.
o terceiro tipo de célula glial é supostamente de origem mesodérmica e, portanto, estritamente falando, não são neurogliais como são os astrocitos e células Muller. Eles entram na retina coincidentes com os precursores mesenquímicos dos vasos sanguíneos retinianos em desenvolvimento (Chan-Ling, 1994). As células microgliais são ubíquas na retina humana sendo encontradas em cada camada da retina.
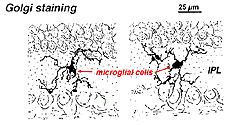
na retina manchada de Golgi parecem formas multipolares estranhas, com corpos celulares pequenos e processos curtos irregulares. De fato, em preparações de Golgi, elas às vezes foram confundidas com células nervosas, particularmente quando elas se encontram em uma camada nuclear com uma única orientação de seus processos na camada plexiforme.Fig. 9. A coloração de Golgi de células microgliais
células microgliais pode ser de dois tipos. Pensa-se que uma forma entra na retina em fases iniciais de desenvolvimento a partir do mesenchyme do nervo óptico e fica dormente nas camadas retinianas durante grande parte da vida da retina. A outra forma de microglia parecem ser células sanguíneas, possíveis provenientes do vaso pericytes (Boycott and Hopkins, 1981; Gallego, 1986). Ambos os tipos podem ser estimulados para uma função macrágica após trauma na retina, e então eles se envolvem em fagocitose de neurônios degeneradores da retina.

Fig. 10a. Lectin-stained microglial cell from Chan-Lin, 1994.
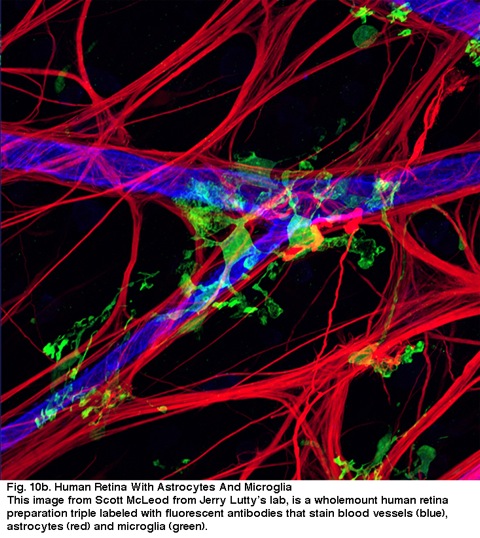
Fig. Retina humana com astrócitos e Microglia. Esta imagem de Scott McLeod do Laboratório de Jerry Lutty, é uma preparação integral de retina humana tripla com anticorpos fluorescentes que mancha vasos sanguíneos (azul), astrócitos (vermelho) e microglia (verde).4. Referência.boicote BB, Hopkins JM. Microglia na retina de macacos e outros mamíferos; sua distinção de outros tipos de glia e células horizontais. Neuro.1981;6:679–688. Cajal SR. In: Thorpe SA, Glickstein M, translators. 1892. A estrutura da retina. Springfield( IL): Thomas; 1972.interacções gliais, neuronais e vasculares na retina dos mamíferos. Programa. Ret. Eye Res. 1994;13:357-389.Edwards RB. Biossíntese do ácido retinóico pelas células glial Müller: um modelo para o sistema nervoso central? Programa. Ret. Eye Res. 1994; 13: 231-242.Fisher SK, Lewis GP. Fotorreceptores e mais além: efeitos celulares e moleculares do descolamento da retina.2nd Great Basin Visual Science Symposium, II, University of Utah Press. 1995
Frishman Lj, Steinberg RH. Aumento provocado pela luz em o na parte proximal da retina de gato adaptada às trevas. J Neurofisiol. 1989;61:1233–1243. Gallego A. estudos comparativos sobre células horizontais e uma nota sobre células microgliais. Programa. Ret. Res. 1986; 5: 165-206.Guerin CJ, Anderson DH, Fisher SK. As alterações na imunolabelagem do filamento intermédio ocorrem em resposta ao descolamento da retina e à recolocação em primatas.Investir. Oftal. Face. Ciência. 1990;31:1474–1482.
Karwoski CJ, Proenza LM. Relação entre as respostas das células Muller, um potencial transretinal local e fluxo de potássio. J Neurofisiol. 1977;40:244–259.Miller RF, Dowling JE. Respostas intracelulares das células Muller (glia) da retina mudpuppy: a sua relação com a onda b do electroretinograma. J Neurofisiol.1970;33:323–341. Newman EA, Odette LL. Model of electroretinogram b-wave generation: a test of the K+ hypothesis. J Neurofisiol. 1984;51:164–182. Newman EA. Electrofisiologia das células gliais da retina. Programa. Ret. Res. 1989; 8: 153-172.Reichenbach A, Robinson SR. O envolvimento das células Müller na retina exterior. In: Djamgoz MBA, Archer SN, Vallerga s, editors. Neurobiologia e aspectos clínicos da retina exterior. London: Chapman & Hall; 1995. P. 395-416.Robinson SR, Hampson ECGM, Munro MN, Vaney DI. Acoplamento unidireccional de entroncamentos entre neuroglia. Procedimento. Austr. Neuroci. Soc. 1993;3:167.Schnitzer J. Astrocytes in mammalian retina. Programa. Ret. Res. 1988; 7: 209-232.Stone J, Makarov F, Hollander H. the glial ensheathment of the soma and axon hillock of retinal ganglion cells. Vis Neurosci. 12:273–279.
Stone J, Dreher Z. relação entre astrócitos, células de gânglios e vasculatura da retina. J Comp Neurol. 1987;255:35–49. Trivino A, Ramirez JM, Salazar JJ, Ramirez AI, Garcia-Sanchez J. Immunohistochemical study of human optic nerve head astroglia. Vision Res. 1996;36:2015-2028. Turner DL, Cepko CL. Um progenitor comum para neurônios e glia persiste na retina do rato no final do desenvolvimento. Natureza. 1987;328:131–136.