pyruvaatkinase M2 brandstoffen meerdere aspecten van kankercellen: van cellulair metabolisme, transcriptionele Regulatie tot extracellulaire signalering
biochemische analyse door karakterisering van de enzymatische activiteit die de vorming van lactaat uit glucose in cellysaten katalyseert onthulde de eerste intracellulaire metabole route, de glycolytische route. Het begin van de zuivering van fracties die glycolytische activiteit bevatten, droeg een aantal pionieronderzoekers bij aan de identificatie van enzymen die in elke stap in de weg impliceren . Deze resultaten bouwen ons moderne concept op in de uitwisseling van aërobe en anaërobe ademhaling en energieproductie onder verschillende fysiologische en pathologische omstandigheden.
het bestaan van een enzym dat de productie van ATP katalyseerde door een fosfaatgroep over te brengen van PEP naar ADP in de lever werd voor het eerst gemeld in 1934 . Daaropvolgende isolatie van het enzym, later bekend als pyruvaatkinase (PK), toonde verschillen aan in weefseldistributie en katalytische kinetiek, wat suggereert dat dit enzym verschillende isovormen kan hebben . Tussen 1982 en 1984 werden verschillende PK-genen gekloond uit gist, kip en rat . De functionele studie van PKM2 werd geïnitieerd door de identificatie van een kandidaatgen in Muis in vroege jaren 1980 . Later, Noguchi et al. toonde aan dat twee isovormen van PK (PKM1 en PKM2) worden gecodeerd door hetzelfde PKM gen via alternatieve splicing . Bij de mens worden PKM-isovormen ook geproduceerd via een soortgelijk splitsingsmechanisme door exon 9 en 10 afzonderlijk op te nemen in pkm1 en pkm2 mRNA .
verschillende bevindingen trokken de aandacht van de onderzoeker op de potentiële rol van PKM2 in tumorigenese. Ten eerste is PKM2 de embryonale isovorm die sterk tot uiting komt tijdens de ontwikkeling van dieren. Zijn transcriptie wordt verzwakt in een aantal volwassen weefsels terwijl het in tumors wordt gereactiveerd . Ten tweede, toonde de studie van de relatieve overvloed van PKM1 en PKM2 in normale en tumorweefsels een omschakeling van isovorm pkm1 aan isovorm PKM2 in diverse kanker zoals hepatocellulair carcinoom aan . Ten derde, wordt de verandering van mRNA het verbinden van PKM1 aan PKM2 verbeterd door c-Myc oncogene die kankercellen voorstellen actief deelnemen aan deze schakelaar om hun vereiste in proliferatie en metabolisme te passen . Ten vierde, modulatie van PKM2 activiteit door activators of inhibitors invloed tumorgroei in vivo .
- de eerste episode: PKM2 als metabolisch enzym in het cytoplasma
- De tweede aflevering: PKM2 als een signalering modulator in het cytoplasma
- de derde episode: PKM2 als transcriptionele regulator in de nucleus
- de vierde episode: PKM2 als extracellulaire signaalcommunicator
- The fifth episode: onbeantwoorde discrepantie van PKM2
- de aanhoudende episode: is PKM2 een kankerbiomarker en drugstreefdoel?
de eerste episode: PKM2 als metabolisch enzym in het cytoplasma
aangezien de rol van PKM2 in metabolische controle van glycolyse in kankercellen uitgebreid is onderzocht , vatten we hier slechts drie cruciale verschillen tussen pkm1 – en PKM2-gemedieerde katalyse en cellulair metabolisme samen. Het eerste verschil is subeenheid interactie. Zowel PKM1 als PKM2 zijn tetramere proteã nen die door vier identieke subeenheden worden gevormd. Elke subeenheid (of monomeer) bevat vier structurele domeinen waaronder A, B, C en n-terminaal domein. Het monomeer dimeriseert eerst samen en dan twee dimeren interageren via de dimeer-dimeer interface georkestreerd door het C-domein van monomeer om een tetramer te vormen. Omdat PKM1 en PKM2 verschillende exons in hun mRNAs omvatten, verandert dit de gecodeerde aminozuren in het domein van C en verandert de tetramerstabiliteit. Onder fysiologische voorwaarde, organiseert pkm1 constitutief als tetrameer terwijl PKM2 in tetrameer of dimeer kan bestaan. Het tweede verschil is allosterische regelgeving. Afhankelijk van de intracellulaire concentraties van kleine molecules en metabolites, kan de activiteit van PKM1 en PKM2 differentieel worden geregeld. Een van de meest bekende allosterische regulatoren is fructose-1,6-bisfosfaat (FBP). Deze glycolytische tussenpersoon bindt PKM2 direct en verhoogt de affiniteit van PKM2 voor PEP . Integendeel, FBP heeft geen significante invloed op de pkm1-activiteit. Naast FBP, zijn andere metabolites, aminozuur en kleine molecules gemeld om pkm2 activiteit te beà nvloeden (Fig. 1). De voor activering of remming vereiste concentratie is echter hoog en het modulerende effect is bescheiden. Of deze kleine moleculen een belangrijke rol spelen in de controle van PKM activiteit onder fysiologische omstandigheden is nog onduidelijk. De activiteit van PKM2 wordt ook geregeld door post-Vertalende wijziging, zoals phosphorylation,acetylation en oxidatie, die de lage activiteit van dimere PKM2 (Fig. 1). Het derde verschil is de energieproduktie en het intermediair gebruik. Aangezien PKM1 constitutief bestaat als het actieve tetrameer, is de belangrijkste biologische functie van deze isovorm de generatie van ATP om cellulaire energie te leveren. Nochtans, kan PKM2, naast ATP produceren, aan de minder actieve dimere vorm overschakelen om verscheidene glycolytic tussenpersonen te produceren die als bouwstenen voor de biosynthese van aminozuren, lipiden en nucleotiden kunnen worden gebruikt.
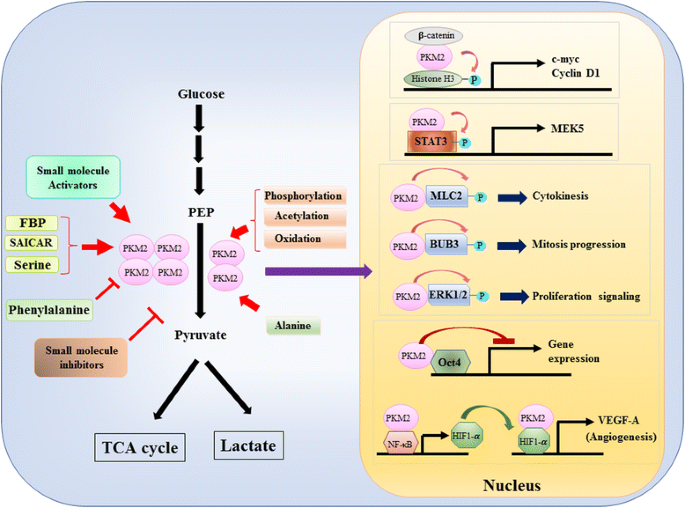
Modulatie van PKM2 activiteit door de fysiologische moleculen en post-translationele modificatie en de potentiële rol van PKM2 in transcriptionele verordening
De tweede aflevering: PKM2 als een signalering modulator in het cytoplasma
naast de functie als glycolytic enzym, PKM2 voorgesteld te betrekken in meer cellulaire processen door de identificatie van de interactie van eiwitten in het cytoplasma. Bijvoorbeeld, PKM2 was er een interactie-eiwit van verschillende tyrosine kinases, waaronder Een-Raf, breakpoint cluster region-Abelson (BCR-ABL) fusie kinase, fibroblast growth factor receptor 1 (FGFR1) enz. . Het is aangetoond dat deze bindingspartners de dimere/tetramere verandering van PKM2 moduleren om het celmetabolisme te veranderen. Het is echter mogelijk dat PKM2 de katalytische kinetiek, de substraatbinding en de cytoplasmatische locatie van deze bindingspartners wederzijds beïnvloedt om de signaaltransductie te moduleren. Het vinden dat PKM2 een fosfor-tyrosine bindende proteã ne is versterkt deze mogelijkheid omdat vele intracellular signalerende bemiddelaars aan fosfor-tyrosineresidu kunnen binden om specifieke eiwitcomplexen voor signaaloverdracht te assembleren . Tot op heden groeit de lijst van pkm2-bindingspartners voortdurend. We belichten verschillende nieuwe leden en bespreken hier hun biologische implicaties. Mukheriee et al. toonde aan dat PKM2 met HuR, een bindende proteã ne van RNA kon binden die een belangrijke rol in de controle van mRNA stabiliteit en Vertalende efficiency speelt, om de vooruitgang van de celcyclus en proliferatie van glioomcellen te bevorderen . Interessant, werd een andere bindende proteã ne tristetraprolin van RNA die een aantal mRNA via het Au-rijke element bij 3′-untranslational gebied (3′-UTR) kon binden ook gevonden om een pkm2 op elkaar inwerkende partner te zijn, en pkm2 veroorzaakte phosphorylation en degradatie van tristetraprolin om de groei van borstkanker te moduleren . Deze twee studies impliceren een potentiële translationele controlefunctie van PKM2. Onlangs, Liang et al. identificeerde het anti-apoptotische eiwit Bcl2 als een nieuwe pkm2 partner . Zij toonden aan dat de oxydatieve spanning de translocatie van PKM2 in mitochondria veroorzaakte waar het gefosforyleerd en bcl2 stabiliseerde door zijn degradatie via ubiquitination-afhankelijke weg te verhinderen. Deze gegevens stelden voor dat pkm2 kankercellen helpt om oxidatieve spanning aan te passen die door intracellular metabolische verandering of extracellulaire belediging wordt opgewekt.
de derde episode: PKM2 als transcriptionele regulator in de nucleus
een nucleaire rol van PKM2 in de regulatie van gentranscriptie of epigenetische modificatie werd in de eerste plaats gesuggereerd door de bevinding dat PKM2 gebonden is met y333-gefosforateerd β-catenine, en het β-catenine-PKM2 complex werd aangetrokken tot de nucleosomen tot fosforylaat Histon H3 bij threonine 11 (Fig. 1). Deze phosphorylation verhoogde later histone H3 acetylation die tot upregulation van β-catenin doelgenen leidde. Een andere transcriptiefactor direct gefosforyleerd door PKM2 is signaaltransducer en activator van transcriptie 3 (STAT3). Pkm2-bemiddelde phosphorylation van STAT3 bij tyrosine 705 verbeterde STAT3 activiteit om de uitdrukking van mitogen-geactiveerde eiwitkinase kinase 5 (MEK5) op te heffen. Naast transcriptiefactoren, is PKM2 getoond aan phosphorylate myosin lichte keten 2 (MLC2), BUB3 en extracellulair signaal-geregeld kinase 1 en 2 (ERK1 en ERK2). Interessant, handelt PKM2 ook via phosphorylation-onafhankelijke manier om genuitdrukking te beà nvloeden. Bijvoorbeeld, is PKM2 gevonden om met Oct4, één van de hoofdtranscriptiefactoren te binden die zelf-vernieuwing van stamcellen controleren, en Oct4-bemiddelde transcriptie remmen . PKM2 kan tumorangiogenese ook verbeteren door met NF-kB en HIF-1α in de kern in wisselwerking te staan en de uitdrukking van het doelgen VEGF-A. van HIF-1α te activeren . Hoewel deze studies sterk de nucleaire localisatie en eiwitkinasefunctie van PKM2 in diverse fysiologische en pathologische omstandigheden suggereerden, nochtans is het belang van nucleaire pkm2-bemiddelde genuitdrukking uitgedaagd door studies gebruikend pkm2 knockoutcellen. Door het gebruik van-geëtiketteerde PEP en PKM2-null muis embryonale fibroblasten, Hosios et al. toonde aan dat PEP-afhankelijke phosphorylation geen gemeenschappelijke gebeurtenis in cellen is en de reactie niet door PKM2 wordt gekatalyseerd . De discrepantie van deze studies is momenteel onopgelost en de eiwitkinaseactiviteit van PKM2 moet verder worden bevestigd.
de vierde episode: PKM2 als extracellulaire signaalcommunicator
de aanwezigheid van extracellulaire PKM2 opent een nieuwe weg voor de studie van de biologische functie van pkm2. Buschow et al. verstrekte het eerste bewijsmateriaal dat PKM2 in B-cel exosomen kon worden ontdekt en als MHC klasse II-geassocieerde proteã ne werd geïdentificeerd . Twee latere studies gaven ook aan dat PKM2 bestaat in exosomes die door diverse cellen worden vrijgegeven . Momenteel, verstrekken verscheidene openbare gegevensbestanden zoals ExoCarta en EVpedia uitvoerige informatie voor de componenten met inbegrip van proteã NEN, lipiden, nucleic zuren van extracellulaire blaasjes in verschillende species. Alle gegevens bevestigen dat PKM2 een pakketeiwit van exosomen is. De recente studies hebben duidelijk een communicatieve rol van exosomes aangetoond door verschillende componenten van gastheercellen aan ontvangende cellen te leveren . Het is te verwachten dat PKM2 een rol kan spelen in cel-cel kruisverwijzing.
nieuw bewijsmateriaal ondersteunt deze hypothese inderdaad. Bijvoorbeeld, toonde een recente studie aan dat bloed doorgevende PKM2 tumorgroei en angiogenese kan bevorderen door de groei, migratie en matrijsadhesie van endothelial cellen te verhogen . Een ander onderzoek toonde ook aan dat PKM2 die van de cellen van de dubbelpuntkanker wordt afgescheiden via een autocrine stimulatie zou kunnen handelen om celmigratie te verbeteren door PI3K/Akt en WNT/β-catenin wegen te activeren . Naast kankercellen, konden neutrofielen bij de plaatsen van de weefselschade PKM2 vrijgeven om angiogenese en het gekronkelde helen te bevorderen . Onze recente studie toonde ook aan dat recombinant pkm2 eiwit fosforylatie en activering van epidermale groei factor receptor (EGFR) kan induceren . Bovendien vonden we dat R339E mutant PKM2 die bij voorkeur dimere PKM2 vormde EGFR meer significant activeerde dan de tetramere PKM2. Keller et al. identificeerde 154 eiwitten als potentiële substraten voor PKM2 na behandeling van Hela-cellen met succinyl-5-aminoimidazol-4-carboxamide-1-ribose-5′ – fosfaat (SAICAR), een intracellulaire metaboliet die de eiwitkinaseactiviteit van PKM2 zou kunnen stimuleren . Ze vonden ook EGFR als PKM2 substraat. Hun resultaten verschillen op twee manieren van die van ons. Ten eerste worden de signalerende routes geactiveerd in onze studie opgewekt door extracellulaire PKM2 terwijl de moleculaire targets geïdentificeerd in hun studie potentiële substraten zijn van intracellulaire PKM2. Ten tweede wordt de toename van ERK1/2-activiteit in onze studie geïnitieerd door EGFR-activering, terwijl erk1/2-activering in hun studie direct wordt gestimuleerd door het saicar/PKM2-complex. Een gelijkaardig fenomeen waargenomen in beide studies is dat r339e mutant PKM2 signaalmoleculen beduidend activeert dan het wilde type PKM2 die de verschillende rol van dimere en tetramere PKM2 in oncogenese suggereren. Door receptortyrosine kinase array te gebruiken, vonden we dat extracellulaire PKM2 slechts beperkte groeifactorreceptoren in borstkankercellen activeerde (gegevens niet getoond). Momenteel is de selectiviteit van receptoractivering door extracellulaire PKM2 onbekend. Bovendien is niet duidelijk waarom R339E mutant PKM2 krachtiger is bij de activering van EGFR. Er zijn meer experimenten nodig om deze vragen te beantwoorden.
een andere elegante vraag die moet worden beantwoord is of vrije PKM2 en blaasjesverpakking PKM2 een vergelijkbaar effect hebben bij het bevorderen van tumorigenese (Fig. 2). Vrije extracellulaire PKM2 kon plasmamembraan niet doordringen en kon intracellular het signaleren via de proteã nen van de celoppervlakte zoals de receptoren van de de groeifactor slechts activeren. Omgekeerd, kon blaasje-verpakkende PKM2 door kanker en stromacellen worden endocytosed, en PKM2 van blaasjes wordt vrijgegeven kon metabolisme en genuitdrukking via intracellular mechanisme beà nvloeden. The PKM2-null cells or mice will be useful to elucidate whether these two extracellular forms of PKM2 could act synergistically or antagonistically in tumorigenesis.

The potential tumor-promoting effect of extracellular PKM2
The fifth episode: onbeantwoorde discrepantie van PKM2
naast de hierboven besproken gegevens op basis van cellen, is de oncogene rol van PKM2 ook uitgedaagd na de generatie van pkm2-knockoutmuizen. Israel et al. gegenereerd een voorwaardelijke knockout muis model door het verwijderen van de pkm2-specifieke exon 10 . Verrassend, versnelde de uitputting van PKM2 maar niet verzwakte tumorvorming die door verlies van BRCA1 gen in muizen wordt gedreven. Uit deze gegevens bleek dat PKM2 niet nodig is voor de proliferatie van kankercellen. Interessant, werd de uitdrukking PKM1 slechts ontdekt in niet-prolifererende tumorcellen die een tumor-onderdrukkende rol van PKM1 in borstkanker suggereren. Bovendien hebben de knockoutmuizen van PKM2 een hoge weerslag om hepatocellulair carcinoom spontaan na een lange latency toe te schrijven aan de onbalans in metabolisme te ontwikkelen . Deze resultaten tegen de notie dat PKM2 een oncogene rol speelt in vivo.
de aanhoudende episode: is PKM2 een kankerbiomarker en drugstreefdoel?
hoewel de resultaten van het genetisch gemanipuleerde muismodel de tumorbevorderende activiteit van PKM2 niet ondersteunen, wordt overexpressie van PKM2 universeel aangetroffen bij menselijke kankers en wordt geassocieerd met een slechte klinische uitkomst (tabel 1) . Twee recente meta-analyses ondersteunden deze conclusie ook. Wu et al. analyseerde de gegevens van 2812 patiënten met stevige tumors van spijsverteringsstelsel die uit 16 cohortstudies worden verkregen en vonden dat de overexpressie van PKM2 met verminderde algemene overleving in maagkanker, slokdarmplaveiselcelcarcinoom, hepatocellular carcinoom, galkanker en mondelinge kanker wordt geassocieerd . PKM2 is echter geen prognostische factor voor alvleesklierkanker. Deze bevinding is niet consistent met eerdere studies waaruit blijkt dat toename van PKM2 een indicator is voor een slechte overleving van pancreaskanker-patiënten . Een ander onderzoek omvatte 4796 gevallen van 27 individuele onderzoeken die aantoonden dat PKM2-upregulatie gecorreleerd is met slechtere totale overleving, ziektevrije overleving en recidiefvrije overleving in gepoolde gegevens . Echter, gestratificeerd door type kanker, voorspelt PKM2 Geen slechte overleving van alvleesklierkanker. Collectief, lijkt PKM2 een aansprakelijk prognostische marker in de meeste stevige tumors te zijn.
integendeel, het gebruik van PKM2 als diagnostische factor is controversieel. Een proteomic analyse toonde aan dat PKM2 een potentiële kenmerkende marker voor de opsporing van longkanker is . Echter, een recente studie suggereerde PKM2 is niet een goede diagnostische marker voor longkanker toe te schrijven aan lage specificiteit . Op dezelfde manier is het onwaarschijnlijk dat PKM2 alleen een nuttige marker is voor de screening van darmkanker . Nochtans, kon de combinatie van veelvoudige markers gevoeligheid en specificiteit voor kankerdiagnose verhogen .
het therapeutisch potentieel van PKM2 is een intrigerende gebeurtenis in de behandeling van kanker. Van de ene kant wordt verwacht dat remming van PKM2 de glycolyse remt, de gentranscriptie belemmert en de cellulaire proliferatie onderdrukt. Daarom lijken pkm2-remmers goede kandidaten te zijn voor de ontwikkeling van geneesmiddelen tegen kanker. Door gebruik te maken van Bibliotheek screening, Vander Heiden et al. identificeerde drie nieuwe klassen van pkm2-inhibitors en toonde aan dat de meest efficiënte samenstelling pkm2-activiteit remde en de dood van kankercellen veroorzaakte . Onlangs hebben Ning et al. gevonden dat nieuwe naftochinonderivaten krachtige pkm2-remmers zijn . Eén effectieve verbinding 3 k onderdrukte de proliferatie van veelvoudige kankercellijnen bij sub-micromolaire concentraties terwijl het weinig schadelijk effect op normale cellen toonde. Van de andere kant, kan de activering van PKM2 tumorgroei ook remmen. Omdat de lage activiteit pkm2 dimeer de belangrijkste isoform is die glycolyse in het cytoplasma en gentranscriptie in de kern in kankercellen teweegbrengt, kunnen pkm2 activatoren die de vorming van tetramere PKM2 kunnen bevorderen glycolyse naar mitochondriënweg schakelen en nucleaire ingang verminderen om gentranscriptie te verzwakken. Beide effecten belemmeren metabolische vraag en de groei-ondersteunende signalering die tot tumorregressie leidt. Twee pioneer-studies identificeerden verschillende pkm2-activatoren en kenmerkten hun specificiteit in vitro . Een volgende studie toonde aan dat pkm2-activatoren inderdaad tetramervorming bevorderden en tumorgroei in vivo onderdrukten . Deze resultaten suggereerden dat pkm2-activatoren veelbelovende anti-kankermedicijnen zouden kunnen zijn.
resistentie tegen chemotherapie is een belangrijke verstopping voor de behandeling van kanker. Het overwinnen van de inherente chemoresistentie van kankercellen is dringend voor klinisch onderzoek. De rollen van PKM2 in chemoresistance van kankercellen zijn geopenbaard, en het richten van PKM2 is getoond om chemoresistant kankercellen opnieuw te sensibiliseren. Een recente studie toonde aan dat CD44 interageert met PKM2 en de pkm2 activiteit onderdrukt via toenemende Tyr105 fosforylatie van PKM2. CD44 ablatie veroorzaakte de omschakeling van aërobe glycolyse naar mitochondriale ademhaling en het verhogen van reactieve zuurstofspecies (ROS) productie, resulterend in de verhoging van cisplatin gevoeligheid in colorectale kankercellen . Remming van de pkm2-activiteit bleek de glycolyse te onderdrukken en cisplatine-resistentie te overwinnen. Daarom kan de combinatie van cisplatine en pkm2-remmers een effectieve strategie zijn voor chemotherapie (Fig. 3). Van PKM2 is ook gemeld dat het bijdraagt aan de regulering van gemcitabine resistentie in pancreaskanker cellen. Kim et al. toonde aan dat pkm2 gemcitabine resistentie bevordert door P38-gemedieerde p53 fosforylering negatief te reguleren, daarom de transcriptionele activiteit van p53 te verminderen en de expressie van pro-apoptotische genen te onderdrukken . Bovendien versterkt ectopische expressie van R399E-PKM2, die bij voorkeur dimere PKM2 vormt, de weerstand van pancreaskanker-cellen tegen gemcitabine. Deze bewijzen suggereren dat PKM2 bijdraagt aan de inductie van geneesmiddelresistentie via een niet-metabolisch mechanisme. Omdat dimere PKM2 hoofdzakelijk als een transcriptionele regulator in kankercellen werkt, kunnen pkm2-activatoren die de vorming van tetramere PKM2 bevorderen effectief zijn in het onderdrukken van de niet-metabolische functie van PKM2 en het verminderen van de weerstand tegen gemcitabine in pancreaskanker cellen. Er is ook aangetoond dat nucleaire PKM2 bijdraagt aan de resistentie van EGFR-remmer bij colorectale kanker en longkanker . Li et al. toonde aan dat de nucleaire pkm2-bemiddelde STAT3 phosphorylation de gevoeligheid van colorectal kankercellen aan gefitinib vermindert en de verstoring van de interactie van nucleaire PKM2 en STAT3 herstelde gefitinibgevoeligheid in de cellen . Meer recent, toonde een andere studie aan dat PKM2 in de kern transloceert en met poly-ADP ribose (PAR) op groeifactor stimulatie in wisselwerking staat. De PAR-bindende activiteit van PKM2 is kritiek voor kernbehoud en gentranscriptie van PKM2 en is belangrijk voor de bevordering van celproliferatie en tumorgroei. Zij concludeerden dat de remming van de kernfunctie van PKM2 de weerstand van EGFR-gemuteerde kankercellen kan overwinnen . Beide studies wezen op een cruciale rol van nucleaire PKM2 in het bemiddelen van geneesmiddelenresistentie bij kanker en suggereerden dat het richten van nucleaire PKM2 een veelbelovende strategie kan zijn om de resistentie te overschrijven.

PKM2 draagt bij tot de proliferatie en geneesmiddelresistentie in kankercellen