piruvat kinaza M2 alimentează mai multe aspecte ale celulelor canceroase: de la metabolismul celular, reglarea transcripțională la semnalizarea extracelulară
analiza biochimică prin caracterizarea activității enzimatice care catalizează formarea lactatului din glucoză în lizatele celulare a relevat prima cale metabolică intracelulară, calea glicolitică. Pornind de la purificarea fracțiunilor care conțineau activitate glicolitică, un număr de cercetători pionieri au contribuit la identificarea enzimelor care implică în fiecare etapă a căii . Aceste rezultate construiesc conceptul nostru modern în schimbul respirației aerobe și anaerobe și al producției de energie în diferite circumstanțe fiziologice și patologice.
existența unei enzime care a catalizat producția de ATP prin transferul unei grupări fosfat de la PEP la ADP în ficat a fost raportată pentru prima dată în 1934 . Izolarea ulterioară a enzimei, cunoscută ulterior sub numele de piruvat kinază (PK), a demonstrat diferențe în distribuția țesuturilor și cinetica catalitică sugerând că această enzimă poate avea izoforme diferite . În perioada 1982-1984, diferite gene PK au fost clonate din drojdie, pui și șobolan . Studiul funcțional al PKM2 a fost inițiat prin identificarea unei gene candidate la șoarece la începutul anilor 1980 . Mai târziu, Noguchi și colab. a arătat că două izoforme ale PK (PKM1 și PKM2) sunt codificate de aceeași genă PKM prin îmbinare alternativă . La om, izoformele PKM sunt, de asemenea, produse printr-un mecanism similar de îmbinare prin includerea exonului 9 și 10 în ARNm PKM1 și pkm2 separat .
Mai multe descoperiri au atras atenția cercetătorului asupra rolului potențial al PKM2 în tumorigeneză. În primul rând, PKM2 este izoforma embrionară care s-a exprimat foarte mult în timpul dezvoltării animalelor. Transcripția sa este atenuată într-un număr de țesuturi adulte în timp ce este reactivată în tumori . În al doilea rând, studiul abundenței relative a PKM1 și PKM2 în țesuturile normale și tumorale a demonstrat o trecere de la izoforma PKM1 la izoforma PKM2 în diferite tipuri de cancer, cum ar fi carcinomul hepatocelular . În al treilea rând, schimbarea îmbinării ARNm de la PKM1 la PKM2 este îmbunătățită de oncogena c-Myc, sugerând că celulele canceroase se angajează activ în acest comutator pentru a se potrivi cerințelor lor în proliferare și metabolism . În al patrulea rând, modularea activității PKM2 de către activatori sau inhibitori afectează creșterea tumorii in vivo .
- primul episod: PKM2 ca enzimă metabolică în citoplasmă
- al doilea episod: pkm2 ca modulator de semnalizare în citoplasmă
- al treilea episod: PKM2 ca regulator transcripțional în nucleul
- al patrulea episod: PKM2 ca comunicator de semnalizare extracelulară
- The fifth episode: discrepanța fără răspuns a pkm2
- episodul continuu: este PKM2 un biomarker de cancer și o țintă de droguri?
primul episod: PKM2 ca enzimă metabolică în citoplasmă
deoarece rolul PKM2 în controlul metabolic al glicolizei în celulele canceroase a fost revizuit pe larg , rezumăm aici doar trei diferențe cruciale între cataliza mediată de PKM1 și PKM2 și metabolismul celular. Prima diferență este interacțiunea subunității. Atât PKM1, cât și PKM2 sunt proteine tetramerice formate din patru subunități identice. Fiecare subunitate (sau monomer) conține patru domenii structurale, inclusiv domeniul A, B, C și n-terminal. Monomerul dimerizează mai întâi împreună și apoi doi dimeri interacționează prin interfața dimer-dimer orchestrată de domeniul C al monomerului pentru a forma un tetramer. Deoarece PKM1 și PKM2 includ exoni diferiți în ARNm-urile lor, Acest lucru modifică aminoacizii codificați din domeniul C și modifică stabilitatea tetramerului. În condiții fiziologice, PKM1 se organizează constitutiv ca tetramer, în timp ce PKM2 poate exista în tetramer sau dimer. A doua diferență este reglarea alosterică. În funcție de concentrațiile intracelulare ale moleculelor mici și ale metaboliților, activitatea PKM1 și PKM2 poate fi reglată diferențiat. Unul dintre cei mai cunoscuți regulatori alosterici este fructoza-1,6-bifosfat (FBP). Acest intermediar glicolitic leagă direct PKM2 și crește afinitatea PKM2 pentru PEP . Dimpotrivă, FBP nu afectează în mod semnificativ activitatea PKM1. În plus față de FBP, s-a raportat că alți metaboliți, aminoacizi și molecule mici afectează activitatea PKM2 (Fig. 1). Cu toate acestea, concentrația necesară pentru activare sau inhibare este ridicată, iar efectul modulator este modest. Dacă aceste molecule mici joacă un rol important în controlul activității PKM în circumstanțe fiziologice este încă neclar. Activitatea PKM2 este, de asemenea, reglementată prin modificări post-translaționale, cum ar fi fosforilarea, acetilarea și oxidarea, care favorizează activitatea scăzută a pkm2 dimeric (Fig. 1). A treia diferență este producția de energie și utilizarea intermediară. Deoarece PKM1 există constitutiv ca tetramer activ, principala funcție biologică a acestei izoforme este generarea de ATP pentru a furniza energie celulară. Cu toate acestea, PKM2, pe lângă producerea ATP, poate trece la forma dimerică mai puțin activă pentru a genera mai mulți intermediari glicolitici care pot fi utilizați ca blocuri de construcție pentru biosinteza aminoacizilor, lipidelor și nucleotidelor.
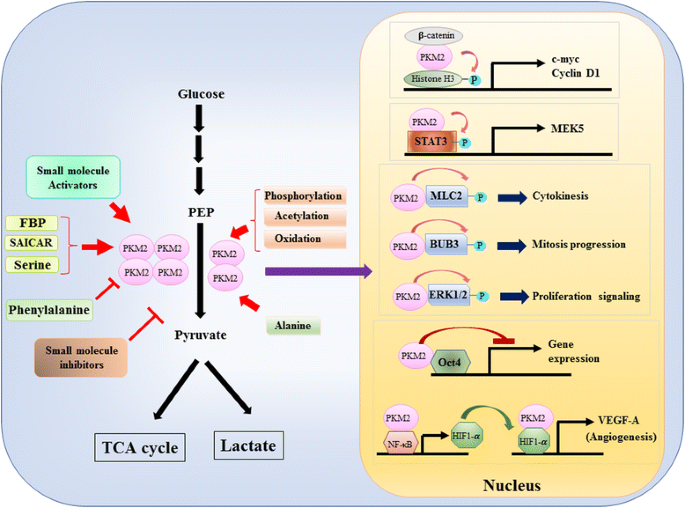
modularea activității PKM2 prin molecule fiziologice și modificări post-translaționale și rolul potențial al PKM2 în reglarea transcripțională
al doilea episod: pkm2 ca modulator de semnalizare în citoplasmă
pe lângă faptul că funcționează ca o enzimă glicolitică, se propune implicarea pkm2 în mai multe procese celulare datorită identificării proteinelor care interacționează în citoplasmă. De exemplu, PKM2 s-a dovedit a fi o proteină care interacționează a mai multor tirozin kinaze, inclusiv A-Raf, Regiunea clusterului punctului de rupere-kinaza de fuziune Abelson (BCR-ABL), receptorul factorului de creștere a fibroblastelor 1 (FGFR1) etc. . S-a demonstrat că acești parteneri de legare modulează modificarea dimerică/tetramerică a PKM2 pentru a modifica metabolismul celular. Cu toate acestea, este posibil ca PKM2 să afecteze reciproc cinetica catalitică, legarea substratului și localizarea citoplasmatică a acestor parteneri de legare pentru a modula transducția semnalului. Constatarea că PKM2 este o proteină de legare a fosforului-tirozinei întărește această posibilitate, deoarece mulți mediatori de semnalizare intracelulară se pot lega de reziduurile de fosfor-tirozină pentru a asambla complexe proteice specifice pentru transmiterea semnalului . Până în prezent, lista partenerilor de legare PKM2 crește continuu. Evidențiem mai mulți membri noi și discutăm implicațiile lor biologice aici. Mukheriee și colab. s-a demonstrat că PKM2 se poate lega cu HuR, o proteină de legare a ARN care joacă un rol important în controlul stabilității ARNm și al eficienței translaționale, pentru a promova progresia ciclului celular și proliferarea celulelor gliomului . Interesant este că o altă proteină de legare a ARN tristetraprolin care ar putea lega un număr de ARNm prin elementul bogat în UA la regiunea 3′-netranslațională (3′-UTR) s-a dovedit, de asemenea, a fi un partener de interacțiune PKM2, iar pkm2 a indus fosforilarea și degradarea tristetraprolinei pentru a modula creșterea cancerului de sân . Aceste două studii implică o potențială funcție de control translațional al PKM2. Recent, Liang și colab. a identificat proteina anti-apoptotică Bcl2 ca un nou partener PKM2 . Ei au demonstrat că stresul oxidativ a indus translocarea PKM2 în mitocondrii unde a fosforilat și stabilizat Bcl2 prin prevenirea degradării sale prin calea dependentă de ubiquitinare. Aceste date au sugerat că PKM2 ajută celulele canceroase să adapteze stresul oxidativ provocat de schimbările metabolice intracelulare sau de insultele extracelulare.
al treilea episod: PKM2 ca regulator transcripțional în nucleul
un rol nuclear al PKM2 în reglarea transcripției genei sau a modificării epigenetice a fost sugerat în primul rând de constatarea că PKM2 legat cu y333-fosforlatizat INQ-catenin, iar complexul INQ-catenin-PKM2 a fost recrutat la nucleozomi pentru a fosforila histona H3 la treonina 11 (Fig. 1). Această fosforilare a crescut ulterior acetilarea histonei H3, ceea ce a dus la reglarea în sus a genelor țintă-catenină. Un alt factor de transcripție fosforilat direct de PKM2 este traductor de semnal și activator al transcripției 3 (STAT3) . Fosforilarea mediată de PKM2 a STAT3 la tirozina 705 a sporit activitatea STAT3 pentru a regla în sus expresia protein kinazei kinazei 5 (MEK5) activată de mitogen. Pe lângă factorii de transcripție, s-a demonstrat că PKM2 fosforilează lanțul ușor de miozină 2 (MLC2), BUB3 și kinaza 1 și 2 reglată de semnal extracelular (ERK1 și ERK2) . Interesant este că PKM2 acționează, de asemenea, printr-o manieră independentă de fosforilare pentru a afecta expresia genelor. De exemplu, s-a constatat că PKM2 se leagă de Oct4, unul dintre factorii de transcripție master care controlează auto-reînnoirea celulelor stem și inhibă transcripția mediată de Oct4 . PKM2 poate spori, de asemenea, angiogeneza tumorală prin interacțiunea cu NF-kB și HIF-1 XV în nucleu și activarea expresiei genei țintă HIF-1 VEGF-A. În consecință, secreția crescută de VEGF-a stimulează formarea vaselor de sânge care contribuie la creșterea tumorii . Deși aceste studii au sugerat cu tărie localizarea nucleară și funcția protein kinazei PKM2 în diferite circumstanțe fiziologice și patologice, cu toate acestea, importanța expresiei genei mediate de pkm2 nuclear a fost contestată de studii folosind celule knockout PKM2. Prin utilizarea-etichetate PEP și pkm2-fibroblaste embrionare de șoarece null, Hosios și colab. a arătat că fosforilarea dependentă de PEP nu este un eveniment comun în celule și reacția nu este catalizată de PKM2 . Discrepanța acestor studii este în prezent nerezolvată, iar activitatea protein kinazei PKM2 necesită o confirmare suplimentară.
al patrulea episod: PKM2 ca comunicator de semnalizare extracelulară
prezența pkm2 extracelular deschide o nouă cale pentru studiul funcției biologice PKM2. Buschow și colab. a furnizat primele dovezi că PKM2 ar putea fi detectat în exozomii celulelor B și a fost identificat ca o proteină asociată clasei MHC II . Două studii ulterioare au indicat, de asemenea, că PKM2 există în exozomii eliberați de diferite celule . În prezent, mai multe baze de date publice precum ExoCarta și EVpedia oferă informații complete pentru componente, inclusiv proteine, lipide, acizi nucleici ai veziculelor extracelulare la diferite specii. Toate datele confirmă faptul că PKM2 este o proteină pachet de exozomi. Studii recente au demonstrat în mod clar un rol comunicativ al exozomilor prin livrarea de componente diferite de la celulele gazdă la celulele receptoare . Este de așteptat ca PKM2 să joace un rol în crosstalk celulă-celulă.
dovezile emergente susțin într-adevăr această ipoteză. De exemplu, un studiu recent a demonstrat că pkm2 care circulă în sânge poate promova creșterea tumorii și angiogeneza prin creșterea creșterii, migrației și aderenței matricei celulelor endoteliale . O altă investigație a arătat, de asemenea, că PKM2 secretat din celulele cancerului de colon ar putea acționa printr-o stimulare autocrină pentru a spori migrarea celulelor prin activarea căilor PI3K/AKT și Wnt/catenin-catenin . În plus față de celulele canceroase, neutrofilele din locurile de deteriorare a țesuturilor ar putea elibera PKM2 pentru a promova angiogeneza și vindecarea rănilor . Studiul nostru recent a demonstrat, de asemenea, că proteina recombinantă PKM2 poate induce fosforilarea și activarea receptorului factorului de creștere epidermal (EGFR) . Mai mult, am constatat că mutantul r339e pkm2 care a format preferențial pkm2 dimeric a activat EGFR mai semnificativ decât pkm2 tetrameric. Keller și colab. au identificat 154 de proteine ca substraturi potențiale pentru PKM2 după tratamentul celulelor Hela cu succinil-5-aminoimidazol-4-carboxamidă-1-riboză-5′-fosfat (SAICAR), un metabolit intracelular care ar putea stimula activitatea protein kinazei PKM2 . De asemenea, au găsit EGFR ca substrat PKM2. Rezultatele lor sunt diferite de ale noastre în două moduri. În primul rând, căile de semnalizare activate în studiul nostru sunt determinate de pkm2 extracelular, în timp ce țintele moleculare identificate în studiul lor sunt substraturi potențiale ale pkm2 intracelular. În al doilea rând, creșterea activității ERK1/2 în studiul nostru este inițiată prin activarea EGFR, în timp ce activarea ERK1/2 în studiul lor este stimulată direct de complexul SAICAR/PKM2. Un fenomen similar observat în ambele studii este că mutantul r339e pkm2 activează moleculele de semnalizare mai semnificativ decât tipul sălbatic PKM2 sugerând rolul distinct al pkm2 dimeric și tetrameric în oncogeneză. Prin utilizarea receptorului tirozin kinazei, am constatat că pkm2 extracelular a activat numai receptorii factorului de creștere limitat în celulele cancerului de sân (datele nu sunt prezentate). În prezent, selectivitatea activării receptorilor de către pkm2 extracelular rămâne necunoscută. În plus, de ce r339e mutant pkm2 este mai puternic în activarea EGFR nu este, de asemenea, clar. Sunt necesare mai multe experimente pentru a răspunde la aceste întrebări.
o altă întrebare elegantă care trebuie abordată este dacă pkm2 liber și ambalajul veziculelor PKM2 prezintă un efect similar în promovarea tumorigenezei (Fig. 2). Pkm2 extracelular liber nu a putut pătrunde în membrana plasmatică și a putut activa doar semnalizarea intracelulară prin intermediul proteinelor de suprafață celulară, cum ar fi receptorii factorului de creștere. În schimb, ambalajul veziculelor PKM2 ar putea fi endocitozat de celulele canceroase și stroma, iar PKM2 eliberat din vezicule ar putea afecta metabolismul și expresia genelor prin mecanismul intracelular. The PKM2-null cells or mice will be useful to elucidate whether these two extracellular forms of PKM2 could act synergistically or antagonistically in tumorigenesis.

The potential tumor-promoting effect of extracellular PKM2
The fifth episode: discrepanța fără răspuns a pkm2
În plus față de datele bazate pe celule discutate mai sus, rolul oncogen al PKM2 a fost, de asemenea, contestat după generarea șoarecilor knockout PKM2. Israelsen și colab. a generat un model de mouse knockout condiționat prin ștergerea exonului specific PKM2 10 . În mod surprinzător, epuizarea PKM2 a accelerat, dar nu a atenuat formarea tumorii determinată de pierderea genei Brca1 la șoareci. Aceste date au indicat faptul că PKM2 nu este necesar pentru proliferarea celulelor canceroase. Interesant este că expresia PKM1 a fost detectată doar în celulele tumorale neproliferante, sugerând un rol supresiv al tumorii PKM1 în cancerul de sân. În plus, șoarecii knockout PKM2 au o incidență ridicată de a dezvolta carcinom hepatocelular spontan după o latență lungă din cauza dezechilibrului metabolismului . Aceste rezultate împotriva noțiunii că PKM2 joacă un rol oncogen in vivo.
episodul continuu: este PKM2 un biomarker de cancer și o țintă de droguri?
deși rezultatele modelului de șoarece de inginerie genetică nu susțin activitatea de promovare a tumorii a PKM2, supraexprimarea PKM2 se găsește universal în cancerele umane și este asociată cu un rezultat clinic slab (Tabelul 1) . Două studii recente de meta-analiză au susținut, de asemenea, această concluzie. Wu și colab. a analizat datele a 2812 pacienți cu tumori solide ale sistemului digestiv obținute din 16 studii de cohortă și a constatat că supraexprimarea PKM2 este asociată cu supraviețuirea globală redusă în cancerul gastric, carcinomul cu celule scuamoase esofagiene, carcinomul hepatocelular, cancerul biliar și cancerul oral . Cu toate acestea, PKM2 nu este un factor prognostic pentru cancerul pancreatic. Această constatare nu este în concordanță cu studiile anterioare care arată că creșterea PKM2 este un indicator al supraviețuirii slabe a pacienților cu cancer pancreatic . O altă investigație a inclus 4796 de cazuri din 27 de studii individuale care au demonstrat că reglarea ascendentă a PKM2 este corelată cu supraviețuirea globală mai slabă, supraviețuirea fără boli și supraviețuirea fără recurență în datele cumulate . Cu toate acestea, stratificat în funcție de tipul de cancer, PKM2 nu prezice o supraviețuire slabă a cancerului pancreatic. În mod colectiv, PKM2 pare a fi un marker prognostic responsabil în majoritatea tumorilor solide.
dimpotrivă, utilizarea PKM2 ca factor de diagnostic este controversată. O analiză proteomică a demonstrat că PKM2 este un marker potențial de diagnostic pentru detectarea cancerului pulmonar . Cu toate acestea, un studiu recent a sugerat că PKM2 nu este un bun marker de diagnostic pentru cancerul pulmonar datorită specificității scăzute . În mod similar, PKM2 singur este puțin probabil să fie un marker util pentru screeningul cancerului de colon . Cu toate acestea, combinația de markeri multipli ar putea crește sensibilitatea și specificitatea pentru diagnosticul de cancer .
potențialul terapeutic al PKM2 este un eveniment interesant în tratamentul cancerului. Pe de o parte, se așteaptă ca inhibarea PKM2 să inhibe glicoliza, să afecteze transcripția genei și să suprime proliferarea celulară. Prin urmare, inhibitorii PKM2 par a fi candidați buni pentru dezvoltarea medicamentelor anti-cancer. Folosind screeningul bibliotecii, Vander Heiden și colab. a identificat trei clase noi de inhibitori PKM2 și a arătat că cel mai eficient compus a inhibat activitatea PKM2 și a indus moartea celulelor canceroase . Recent, Ning și colab. s-a constatat că derivații noi de naftochinonă sunt inhibitori puternici ai PKM2 . Un compus eficient 3 k a suprimat proliferarea mai multor linii celulare canceroase la concentrații sub-micromolare, în timp ce a arătat un efect dăunător redus asupra celulelor normale. Pe de altă parte, activarea PKM2 poate inhiba, de asemenea, creșterea tumorii. Deoarece dimerul pkm2 cu activitate scăzută este izoforma majoră care declanșează glicoliza în citoplasmă și transcripția genică în nucleu în celulele canceroase, activatorii pkm2 care pot promova formarea pkm2 tetramerici pot comuta glicoliza pe calea mitocondriilor și pot reduce intrarea nucleară pentru a atenua transcripția genei. Ambele efecte afectează cererea metabolică și semnalizarea de susținere a creșterii care duce la regresia tumorii. Două studii de pionierat au identificat diferiți activatori PKM2 și au caracterizat specificitatea lor in vitro . Un studiu ulterior a demonstrat că activatorii PKM2 au promovat într-adevăr formarea tetramerului și au suprimat creșterea tumorii in vivo . Aceste rezultate au sugerat că activatorii PKM2 ar putea promite medicamente anti-cancer.
rezistența la chimioterapie este un blocaj major pentru tratamentul cancerului. Depășirea chimiorezistenței inerente a celulelor canceroase este urgentă pentru cercetarea clinică. Rolurile PKM2 în chimiorezistența celulelor canceroase au fost dezvăluite, iar direcționarea PKM2 s-a dovedit a re-sensibiliza celulele canceroase chimiorezistente. Un studiu recent a arătat că CD44 interacționează cu PKM2 și suprimă activitatea PKM2 prin creșterea fosforilării Tyr105 a PKM2. Ablația CD44 a indus trecerea de la glicoliza aerobă la respirația mitocondrială și creșterea producției de specii reactive de oxigen (ROS), rezultând creșterea sensibilității cisplatinei în celulele canceroase colorectale . S-a demonstrat că inhibarea activității PKM2 suprimă glicoliza și depășește rezistența la cisplatină. Prin urmare, combinația de cisplatină și inhibitori ai PKM2 poate fi o strategie eficientă pentru chimioterapie (Fig. 3). De asemenea, s-a raportat că PKM2 participă la reglarea rezistenței la gemcitabină în celulele canceroase pancreatice. Kim și colab. a arătat că PKM2 promovează rezistența la gemcitabină prin reglarea negativă a fosforilării P38 mediate de p53, reducând astfel activitatea transcripțională a p53 și suprimând expresia genelor pro-apoptotice . Mai mult, expresia ectopică a R399E-PKM2, care formează preferențial pkm2 dimeric, îmbunătățește rezistența celulelor canceroase pancreatice la gemcitabină. Aceste dovezi sugerează că PKM2 contribuie la inducerea rezistenței la medicamente printr-un mecanism non-metabolic. Deoarece pkm2 dimeric acționează în principal ca un regulator transcripțional în celulele canceroase, activatorii PKM2 care promovează formarea pkm2 tetrameric pot fi eficienți în suprimarea funcției non-metabolice a PKM2 și reducerea rezistenței la gemcitabină în celulele canceroase pancreatice. S-a demonstrat, de asemenea, că pkm2 Nuclear contribuie la rezistența inhibitorului EGFR în cancerul colorectal și cancerul pulmonar . Li și colab. a arătat că fosforilarea STAT3 mediată de pkm2 nuclear reduce sensibilitatea celulelor canceroase colorectale la gefitinib și întreruperea interacțiunii dintre pkm2 nuclear și STAT3 restaurat sensibilitatea gefitinib în celule . Mai recent, un alt studiu a demonstrat că PKM2 se translocă în nucleu și interacționează cu riboza poli-ADP (PAR) la stimularea factorului de creștere. Activitatea de legare PAR a PKM2 este critică pentru retenția nucleară și transcripția genică a PKM2 și este importantă pentru promovarea proliferării celulare și a creșterii tumorale. Ei au concluzionat că inhibarea funcției nucleare PKM2 poate depăși rezistența celulelor canceroase mutante EGFR . Ambele studii au subliniat un rol crucial al pkm2 nuclear în medierea rezistenței la medicamente în cancere și au sugerat că direcționarea pkm2 nuclear poate fi o strategie promițătoare pentru a depăși rezistența.

PKM2 contribuie la proliferarea și rezistența la medicamente în celulele canceroase