pyruvatkinas M2 bränner flera aspekter av cancerceller: från cellulär metabolism, transkriptionsreglering till extracellulär signalering
Biokemisk analys genom att karakterisera den enzymatiska aktiviteten som katalyserar bildandet av laktat från glukos i celllysater avslöjade den första intracellulära metaboliska vägen, den glykolytiska vägen. Från och med rening av fraktioner som innehöll glykolytisk aktivitet bidrog ett antal pionjärforskare till identifieringen av enzymer som involverar i varje steg i vägen . Dessa resultat bygger upp vårt moderna koncept i utbyte av aerob och anaerob andning och energiproduktion under olika fysiologiska och patologiska omständigheter.
förekomsten av ett enzym som katalyserade produktionen av ATP genom att överföra en fosfatgrupp från PEP till ADP i levern rapporterades först 1934 . Efterföljande isolering av enzymet, känt som pyruvatkinas (PK) senare, visade skillnader i vävnadsfördelning och katalytisk kinetik vilket tyder på att detta enzym kan ha olika isoformer . Under 1982 till 1984 klonades olika PK-gener från jäst, kyckling och råtta . Den funktionella studien av PKM2 initierades genom identifiering av en kandidatgen i mus i början av 1980-talet . Senare, Noguchi et al. visade att två isoformer av PK (PKM1 och PKM2) kodas av samma PKM-gen via Alternativ Splitsning . Hos människa produceras PKM-isoformer också via en liknande skarvningsmekanism genom att inkludera exon 9 och 10 i PKM1 och PKM2 mRNA separat .
flera fynd fångade forskarens uppmärksamhet på den potentiella rollen av PKM2 i tumörgenes. För det första är PKM2 den embryonala isoformen som starkt uttrycks under djurutveckling. Dess transkription dämpas i ett antal vuxna vävnader medan den återaktiveras i tumörer . För det andra visade studien av det relativa överflödet av PKM1 och PKM2 i normala och tumörvävnader en övergång från pkm1-isoformen till PKM2-isoformen i olika cancerformer som hepatocellulärt karcinom . För det tredje förbättras förändringen av mRNA-skarvning från PKM1 till PKM2 av c-Myc-onkogen som tyder på att cancerceller aktivt engagerar sig i denna omkopplare för att passa deras krav på proliferation och metabolism . För det fjärde påverkar modulering av PKM2-aktivitet av aktivatorer eller hämmare tumörtillväxt in vivo .
- första avsnittet: PKM2 som ett metaboliskt enzym i cytoplasman
- det andra avsnittet: pkm2 som en signalmodulator i cytoplasman
- det tredje avsnittet: PKM2 som en transkriptionsregulator i kärnan
- den fjärde episoden: PKM2 som en extracellulär signalkommunikator
- The fifth episode: obesvarad skillnad mellan PKM2
- den fortsatta episoden: är PKM2 en cancerbiomarkör och läkemedelsmål?
första avsnittet: PKM2 som ett metaboliskt enzym i cytoplasman
eftersom pkm2: s roll i metabolisk kontroll av glykolys i cancerceller har granskats omfattande , sammanfattar vi bara tre avgörande skillnader mellan PKM1 – och PKM2-medierad katalys och cellulär metabolism här. Den första skillnaden är subenhetsinteraktion. Både PKM1 och PKM2 är tetramera proteiner bildade av fyra identiska underenheter. Varje underenhet (eller monomer) innehåller fyra strukturella domäner inklusive A, B, C och N-terminaldomän. Monomeren dimeriserar först tillsammans och sedan interagerar två dimerer via dimer-dimer-gränssnittet orkestrerat av C-domänen för monomer för att bilda en tetramer. Eftersom PKM1 och PKM2 inkluderar olika exoner i deras mRNA, förändrar detta de kodade aminosyrorna i C-domänen och förändrar tetramerstabiliteten. Under fysiologiskt tillstånd organiserar PKM1 konstitutivt som en tetramer medan PKM2 kan existera i tetramer eller dimer. Den andra skillnaden är allosterisk reglering. Beroende på de intracellulära koncentrationerna av små molekyler och metaboliter kan aktiviteten hos PKM1 och PKM2 regleras differentiellt. En av de mest kända allosteriska regulatorerna är fruktos-1,6-bisfosfat (FBP). Denna glykolytiska mellanprodukt binder direkt PKM2 och ökar affiniteten hos PKM2 för PEP . Tvärtom påverkar FBP inte signifikant pkm1-aktivitet. Förutom FBP har andra metaboliter, aminosyra och små molekyler rapporterats påverka pkm2-aktivitet (Fig. 1). Koncentrationen som krävs för aktivering eller hämning är emellertid hög och den modulerande effekten är blygsam. Huruvida dessa små molekyler spelar en viktig roll i kontrollen av PKM-aktivitet under fysiologiska omständigheter är fortfarande oklart. PKM2-aktivitet regleras också genom post-translationell modifiering, såsom fosforylering, acetylering och oxidation, vilket gynnar den låga aktiviteten hos dimerisk PKM2 (Fig. 1). Den tredje skillnaden är energiproduktion och mellanliggande användning. Eftersom pkm1 konstitutivt existerar som den aktiva tetrameren är den huvudsakliga biologiska funktionen hos denna isoform genereringen av ATP för att leverera cellulär energi. Emellertid kan PKM2, förutom att producera ATP, byta till den mindre aktiva dimera formen för att generera flera glykolytiska mellanprodukter som kan användas som byggstenar för biosyntes av aminosyror, lipider och nukleotider.
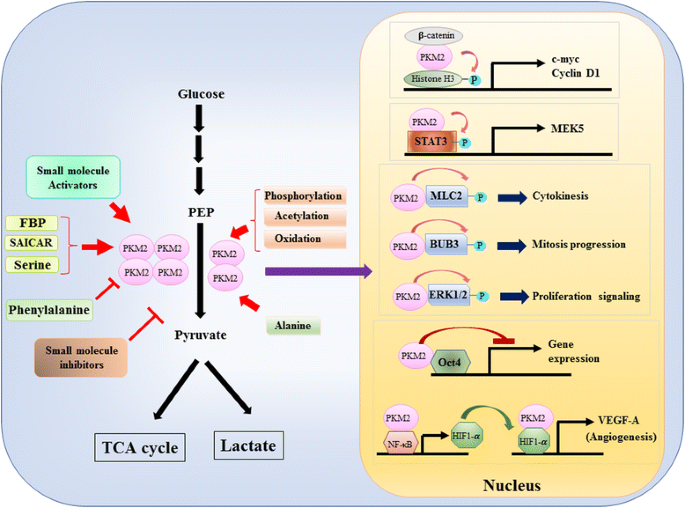
modulering av pkm2-aktivitet genom fysiologiska molekyler och post-translationell modifiering och pkm2: s potentiella roll i transkriptionsreglering
det andra avsnittet: pkm2 som en signalmodulator i cytoplasman
förutom att fungera som ett glykolytiskt enzym föreslås pkm2 att involvera i mer cellulära processer på grund av identifieringen av interagerande proteiner i cytoplasman. Till exempel visades PKM2 vara ett interagerande protein av flera tyrosinkinaser inklusive A-Raf, Brytpunktsklusterregion-Abelson (BCR-ABL) fusionskinas, fibroblasttillväxtfaktorreceptor 1 (FGFR1) etc. . Dessa bindningspartners har visat sig modulera den dimera / tetramera förändringen av PKM2 för att förändra cellmetabolismen. Det är emellertid möjligt att PKM2 ömsesidigt kan påverka katalytisk kinetik, substratbindning och cytoplasmatisk placering av dessa bindningspartners för att modulera signaltransduktion. Upptäckten att PKM2 är ett fosfortyrosinbindande protein stärker denna möjlighet eftersom många intracellulära signalmediatorer kan binda till fosfortyrosinrester för att montera specifika proteinkomplex för signalöverföring . Hittills växer listan över pkm2-bindande partners kontinuerligt. Vi lyfter fram flera nya medlemmar och diskuterar deras biologiska konsekvenser här. Mukheriee et al. demonstrerade att PKM2 kunde binda med HuR, ett RNA-bindande protein som spelar en viktig roll i kontrollen av mRNA-stabilitet och translationell effektivitet, för att främja cellcykelprogression och proliferation av gliomceller . Intressant, en annan RNA-bindande protein tristetraprolin som kunde binda ett antal mRNA via AU-rika elementet vid 3′-untranslational region (3′ – UTR) befanns också vara en pkm2 samverkande partner, och PKM2 inducerad fosforylering och nedbrytning av tristetraprolin att modulera bröstcancertillväxt . Dessa två studier innebär en potentiell translationell kontrollfunktion av PKM2. Nyligen, Liang et al. identifierade det anti-apoptotiska proteinet Bcl2 som en ny pkm2-partner . De visade att oxidativ stress inducerade translokationen av PKM2 till mitokondrier där den fosforylerade och stabiliserade Bcl2 genom att förhindra dess nedbrytning via ubiquitinationsberoende väg. Dessa data föreslog att PKM2 hjälper cancerceller att anpassa oxidativ stress som framkallas av intracellulär metabolisk förändring eller extracellulär förolämpning.
det tredje avsnittet: PKM2 som en transkriptionsregulator i kärnan
en nukleär roll av PKM2 i regleringen av gentranskription eller epigenetisk modifiering föreslogs först av upptäckten att PKM2 bunden med y333-fosforlaterad Bisexuell-catenin, och komplexet för att rekrytera till nukleosomerna för att fosforylera Histon H3 vid treonin 11 (Fig. 1). Denna fosforylering ökade därefter Histon H3-acetylering som ledde till uppreglering av målgener från målgruppen för mål. En annan transkriptionsfaktor som direkt fosforyleras av PKM2 är signalomvandlare och aktivator för transkription 3 (STAT3) . PKM2-medierad fosforylering av STAT3 vid tyrosin 705 förbättrad STAT3-aktivitet för att uppreglera uttrycket av mitogenaktiverat proteinkinas Kinas 5 (MEK5). Förutom transkriptionsfaktorer har PKM2 visat sig fosforylera myosinljuskedjan 2 (MLC2), BUB3 och extracellulärt signalreglerat Kinas 1 och 2 (ERK1 och ERK2) . Intressant verkar PKM2 också via fosforyleringsoberoende sätt att påverka genuttryck. Till exempel har PKM2 befunnits binda med Oct4, en av huvudtranskriptionsfaktorerna som kontrollerar självförnyelse av stamceller och hämmar Oct4-medierad transkription . PKM2 kan också förbättra tumörangiogenesen genom att interagera med NF-kB och HIF-1 kg i kärnan och aktivera uttrycket av HIF-1 kg målgen VEGF-A. följaktligen ökar ökad utsöndring av VEGF-A blodkärlbildning som bidrar till tumörtillväxt . Även om dessa studier starkt föreslog kärnlokalisering och proteinkinasfunktion av PKM2 i olika fysiologiska och patologiska omständigheter, men vikten av nukleärt PKM2-medierat genuttryck har utmanats av studier med pkm2 knockout-celler. Genom att använda-märkta PEP-och PKM2-null-mus embryonala fibroblaster, Hosios et al. visade att PEP-beroende fosforylering inte är en vanlig händelse i celler och reaktionen katalyseras inte av PKM2 . Skillnaden mellan dessa studier är för närvarande olöst och proteinkinasaktiviteten hos PKM2 behöver ytterligare bekräftelse.
den fjärde episoden: PKM2 som en extracellulär signalkommunikator
närvaron av extracellulär PKM2 öppnar en ny aveny för studier av pkm2 biologisk funktion. Buschow et al. tillhandahöll det första beviset att PKM2 kunde detekteras i B-cellsexosomer och identifierades som ett MHC-klass II-associerat protein . Två efterföljande studier indikerade också att PKM2 fanns i exosomer som släpptes av olika celler . För närvarande ger flera offentliga databaser som ExoCarta och EVpedia omfattande information för komponenterna inklusive proteiner, lipider, nukleinsyror av extracellulära vesiklar i olika arter. Alla data bekräftar att PKM2 är ett paketprotein av exosomer. Nya studier har tydligt visat en kommunikativ roll av exosomer genom att leverera olika komponenter från värdceller till mottagarceller . Det kan förväntas att PKM2 kan spela en roll i cellcellöverhörning.
nya bevis stöder verkligen denna hypotes. Till exempel visade en ny studie att blodcirkulerande PKM2 kan främja tumörtillväxt och angiogenes genom att öka tillväxten, migrationen och matrisadhesionen av endotelceller . En annan undersökning visade också att PKM2 utsöndrat från koloncancerceller kan verka via en autokrin stimulering för att förbättra cellmigrering genom att aktivera PI3K/Akt och Wnt/Baccarat-catenin vägar . Förutom cancerceller kan neutrofiler vid vävnadsskadorna frigöra PKM2 för att främja angiogenes och sårläkning . Vår senaste studie visade också att rekombinant pkm2-protein kan inducera fosforylering och aktivering av epidermal tillväxtfaktorreceptor (EGFR) . Dessutom fann vi att r339e mutant PKM2 som företrädesvis bildade dimerisk PKM2 aktiverade EGFR mer signifikant än den tetrameriska PKM2. Keller et al. identifierade 154 proteiner som potentiella substrat för PKM2 efter behandling av Hela-celler med succinyl-5-aminoimidazol-4-karboxamid-1-ribos-5′ – fosfat (SAICAR), en intracellulär metabolit som kan stimulera proteinkinasaktiviteten hos PKM2 . De fann också EGFR som ett pkm2-substrat. Deras resultat skiljer sig från våra på två sätt. Först framkallas signalvägarna som aktiveras i vår studie av extracellulär PKM2 medan de molekylära målen som identifierats i deras studie är potentiella substrat av intracellulär PKM2. För det andra initieras ökning av erk1/2-aktivitet i vår studie genom EGFR-aktivering medan erk1/2-aktivering i deras studie stimuleras direkt av SAICAR/PKM2-komplexet. Ett liknande fenomen som observerats i båda studierna är att r339e mutant PKM2 aktiverar signalmolekyler mer signifikant än vildtypen PKM2 vilket tyder på den distinkta rollen av dimerisk och tetramerisk PKM2 i onkogenes. Genom att använda receptortyrosinkinasuppsättning fann vi att extracellulär PKM2 endast aktiverade begränsade tillväxtfaktorreceptorer i bröstcancerceller (data visas inte). För närvarande är selektiviteten för receptoraktivering av extracellulär PKM2 fortfarande okänd. Dessutom, varför R339E mutant PKM2 är mer potent i aktiveringen av EGFR är inte heller klart. Fler experiment behövs för att svara på dessa frågor.
en annan elegant fråga som ska behandlas är om fri PKM2 och vesikelförpacknings PKM2 uppvisar liknande effekt för att främja tumörgenes (Fig. 2). Fri extracellulär PKM2 kunde inte tränga igenom plasmamembranet och kunde bara aktivera intracellulär signalering via cellytproteiner som tillväxtfaktorreceptorer. Omvänt kan vesikelförpackningspkm2 endocytoseras av cancer-och stromaceller, och PKM2 som frigörs från vesiklar kan påverka metabolism och genuttryck via intracellulär mekanism. The PKM2-null cells or mice will be useful to elucidate whether these two extracellular forms of PKM2 could act synergistically or antagonistically in tumorigenesis.

The potential tumor-promoting effect of extracellular PKM2
The fifth episode: obesvarad skillnad mellan PKM2
förutom de cellbaserade data som diskuterats ovan har den onkogena rollen för PKM2 också utmanats efter genereringen av pkm2 knockout-möss. Israelsen et al. genererade en villkorlig knockout-musmodell genom att radera den PKM2-specifika exon 10 . Överraskande accelererade utarmningen av PKM2 men inte dämpad tumörbildning som drivs av förlust av Brca1-genen hos möss. Dessa data indikerade att PKM2 inte krävs för proliferation av cancerceller. Intressant nog detekterades pkm1-uttryck endast i icke-prolifererande tumörceller som tyder på en tumörundertryckande roll av PKM1 i bröstcancer. Dessutom har pkm2 knockout-möss en hög förekomst att utveckla hepatocellulärt karcinom spontant efter en lång latens på grund av obalansen i ämnesomsättningen . Dessa resultat mot uppfattningen att PKM2 spelar en onkogen roll in vivo.
den fortsatta episoden: är PKM2 en cancerbiomarkör och läkemedelsmål?
även om resultaten av genetiskt ingenjörsmässig musmodell inte stöder den tumörfrämjande aktiviteten hos PKM2, finns överuttryck av PKM2 universellt i humana cancerformer och är associerad med dåligt kliniskt resultat (tabell 1) . Två nya metaanalysstudier stödde också denna slutsats. Wu et al. analyserade data från 2812 patienter med fasta tumörer i matsmältningssystemet som erhållits från 16 kohortstudier och fann att överuttryck av PKM2 är associerad med minskad total överlevnad i magcancer, esofagus skivepitelcancer, hepatocellulärt karcinom, gallcancer och oral cancer . PKM2 är dock inte en prognostisk faktor för bukspottkörtelcancer. Detta resultat överensstämmer inte med tidigare studier som visar att ökning av PKM2 är en indikator på dålig överlevnad hos patienter med bukspottkörtelcancer . En annan undersökning inkluderade 4796 fall från 27 enskilda studier visade att PKM2-uppreglering är korrelerad med sämre total överlevnad, sjukdomsfri överlevnad och återfallsfri överlevnad i poolade data . Men stratifierad av cancertyp förutsäger PKM2 inte en dålig överlevnad av bukspottskörtelcancer. Sammantaget verkar PKM2 vara en ansvarig prognostisk markör i de flesta fasta tumörer.
tvärtom är användningen av PKM2 som diagnostisk faktor kontroversiell. En proteomisk analys visade att PKM2 är en potentiell diagnostisk markör för detektering av lungcancer . En ny studie föreslog dock att PKM2 inte är en bra diagnostisk markör för lungcancer på grund av låg specificitet . På samma sätt är PKM2 ensam osannolikt att vara en användbar markör för screening av koloncancer . Kombination av flera markörer kan dock öka känsligheten och specificiteten för cancerdiagnos .
den terapeutiska potentialen hos PKM2 är en spännande händelse i cancerbehandling. Från ena sidan förväntas hämning av PKM2 hämma glykolys, försämra gentranskription och undertrycka cellulär proliferation. Därför verkar PKM2-hämmare vara bra kandidater för utveckling av cancer mot cancer. Genom att använda biblioteksscreening, Vander Heiden et al. identifierade tre nya klasser av pkm2-hämmare och visade att den mest effektiva föreningen hämmade PKM2-aktivitet och inducerade död av cancerceller . Nyligen, Ning et al. fann att nya naftokinonderivat är potenta PKM2-hämmare . En effektiv förening 3 k undertryckte proliferationen av flera cancercellinjer vid submikromolära koncentrationer medan den visade liten skadlig effekt på normala celler. Från den andra sidan kan aktivering av PKM2 också hämma tumörtillväxt. Eftersom den låga aktiviteten PKM2-dimeren är den huvudsakliga isoformen som utlöser glykolys i cytoplasman och gentranskription i kärnan i cancerceller, kan PKM2-aktivatorer som kan främja bildandet av tetramerisk PKM2 byta glykolys till mitokondrierväg och minska kärninträde för att dämpa gentranskription. Båda effekterna försämrar metabolisk efterfrågan och tillväxtstödjande signalering som leder till tumörregression. Två pionjärstudier identifierade olika pkm2-aktivatorer och karakteriserade deras specificitet in vitro . En efterföljande studie visade att PKM2-aktivatorer verkligen främjade tetramerbildning och undertryckte tumörtillväxt in vivo . Dessa resultat föreslog PKM2-aktivatorer kan vara lovande läkemedel mot cancer.
resistens mot kemoterapi är en stor blockering för cancerbehandling. Att övervinna den inneboende kemoresistansen hos cancerceller är brådskande för klinisk forskning. Rollerna av PKM2 i chemoresistance av cancerceller har avslöjats, och uppsätta som mål PKM2 har visats för att re-sensibilisera chemoresistant cancerceller. En ny studie visade att CD44 interagerar med PKM2 och undertrycker PKM2-aktivitet via ökande tyr105-fosforylering av PKM2. CD44-ablation inducerade övergången från aerob glykolys till mitokondriell andning och ökad produktion av reaktiva syrearter (ROS), vilket resulterade i förbättring av cisplatinkänslighet i kolorektala cancerceller . Hämning av pkm2-aktivitet visades undertrycka glykolys och övervinna cisplatinresistens. Därför kan kombinationen av cisplatin-och PKM2-hämmare vara en effektiv strategi för kemoterapi (Fig. 3). PKM2 har också rapporterats delta i regleringen av gemcitabinresistens i bukspottkörtelcancerceller. Kim et al. visade att PKM2 främjar gemcitabinresistens genom att negativt reglera p38-medierad p53-fosforylering, vilket minskar transkriptionsaktiviteten hos p53 och undertrycker uttrycket av pro-apoptotiska gener . Dessutom ökar ektopiskt uttryck av R399E-PKM2, som företrädesvis bildar dimerisk PKM2, resistensen hos bukspottkörtelcancerceller mot gemcitabin. Dessa bevis tyder på att PKM2 bidrar till induktion av läkemedelsresistens via en icke-metabolisk mekanism. Eftersom dimerisk PKM2 huvudsakligen fungerar som en transkriptionsregulator i cancerceller, kan PKM2-aktivatorer som främjar bildandet av tetramerisk PKM2 vara effektiva för att undertrycka icke-metabolisk funktion av PKM2 och minska resistensen mot gemcitabin i bukspottkörtelcancerceller. Nukleär PKM2 har också visats bidra till resistens hos EGFR-hämmare vid kolorektal cancer och lungcancer . Li et al. visade att kärn PKM2-medierad STAT3 fosforylering minskar känsligheten hos kolorektala cancerceller till gefitinib och störning av interaktionen av kärn PKM2 och STAT3 återställd gefitinib känslighet i cellerna . På senare tid visade en annan studie att PKM2 translokerar in i kärnan och interagerar med poly-ADP-ribos (PAR) vid tillväxtfaktorstimulering. Den PARBINDANDE aktiviteten hos PKM2 är kritisk för kärnretention och gentranskription av PKM2 och är viktig för främjande av cellproliferation och tumörtillväxt. De drog slutsatsen att hämning av pkm2-kärnfunktion kan övervinna motståndet hos EGFR-muterade cancerceller . Båda studierna påpekade en avgörande roll för nukleär PKM2 vid förmedling av läkemedelsresistens i cancer och föreslog att inriktning på nukleär PKM2 kan vara en lovande strategi för att åsidosätta motståndet.

PKM2 bidrar till proliferation och läkemedelsresistens i cancerceller