Vitamin C: ett antioxidantmedel
Redoxmetabolism och antioxidantegenskaper hos vitamin C
fria radikaler och oxidanter spelar en dubbel roll som både toxiska och fördelaktiga föreningar, i metaboliska processer och som svar på exogena stimuleringar. De produceras antingen från normala metaboliska aktiviteter eller från miljöfaktorer (förorening, cigarettrök och strålning). När en överbelastning av fria radikaler inte kan rensas, genererar deras ackumulering i kroppen oxidativ stress . Oxidativ stress uppstår när friradikalbildning överstiger förmågan att skydda mot dem. Denna process leder till utveckling av kroniska och degenerativa sjukdomar som cancer, autoimmuna störningar, åldrande, katarakt, reumatoid artrit, hjärt-och neurodegenerativa sjukdomar . En antioxidant är en molekyl som förhindrar oxidation av andra molekyler. Oxidationsprocessen är en kemisk reaktion som producerar fria radikaler, vilket leder till kedjereaktioner som kan skada celler. Antioxidanteffekten av C-vitamin har dokumenterats väl . C-Vitamin är en kraftfull antioxidant som har förmåga att donera en väteatom och bilda en relativt stabil askorbylfri radikal. E-Vitamin, C-vitamin och karotenkaroten är kända som antioxidantvitaminer som föreslås minska oxidativ skada och sänka risken för vissa kroniska sjukdomar. Sjukdomar, såsom hjärt-och kärlsjukdomar, är förknippade med otillräckliga koncentrationer av l-askorbinsyra, tokoferol och sackarotenkaroten i epidemiologiska studier. C-Vitamin ökar också järnabsorptionen genom att minska Fe3+ till Fe2+ från icke-hemjärnkällor . I närvaro av redoxaktiva joner (järn, koppar) fungerar C-vitamin som en prooxidant, vilket bidrar till bildandet av hydroxylradikaler, vilket kan leda till lipid -, DNA-eller proteinoxidation . Det finns olika mekanismer för att lindra oxidativ stress och reparera skadade makromolekyler. Enzymatiska och icke-enzymatiska antioxidanter har viktiga roller i avlägsnande av fria radikaler och reaktiva syrearter (ROS). Antioxidantenzymerna, katalas (CAT), superoxiddismutas (SOD), glutationreduktas (GR), glutationperoxidas (GSHpx) och, i växter, askorbatperoxidas (AA-px) och de icke-enzymatiska antioxidanterna, inklusive glutation (GSH) och askorbat (ASC), har visat sig påverkas signifikant av oxidativ stress . Antioxidantföreningar kan förhindra okontrollerad bildning av fria radikaler eller hämma deras reaktion med biologiska platser; även förstörelsen av de flesta fria radikaler beror på oxidationen av endogena antioxidanter, huvudsakligen genom att avlägsna och reducera molekyler . C-Vitamin anses vara en viktig vattenlöslig antioxidant som rapporteras neutralisera ROS och minska oxidativ stress .
C-Vitamin är ett potent reduktionsmedel och rensare av fria radikaler i biologiska system . Det är involverat i den första raden av antioxidantförsvar, skyddar lipidmembran och proteiner från oxidativ skada. Som en vattenlöslig molekyl kan C-vitamin fungera både inom och utanför cellerna och kan neutralisera fria radikaler och förhindra skador på fria radikaler. C-Vitamin är en utmärkt källa till elektroner för fria radikaler som söker en elektron för att återfå sin stabilitet. C-Vitamin kan donera elektroner till fria radikaler och släcka deras reaktivitet .
C-Vitamin har visat sig vara en effektiv rensare mot syre-och kväveoxidarter, såsom superoxidradikaljon, väteperoxid, hydroxylradikalen och singlet syre. Denna egenskap av C-vitamin har viktiga processer för att skydda cellulära komponenter från friradikalinducerad skada. Dessutom är C-vitamin effektivt för att regenerera antioxidantformen av E-vitamin genom att minska tokoferoxylradikalerna. Denna process skyddar membran och andra fack i cellen från fria radikaler-inducerad skada (Figur 2). Askorbatperoxidas (APX) är ett enzym som reducerar H2O2 till vatten genom att använda askorbat som elektrondonator. Monodehydroascorbat är ett oxiderat askorbat som regenereras av monodehydroascorbatreduktas (MDAR). Monodehydroaskorbatradikal disproportionerar snabbt till askorbat och dehydroaskorbat. Dehydroascorbate is reduced to ascorbate by dehydroascorbate reductase in the presence of GSH, yielding oxidized glutathione (GSSG). It is reduced by glutathione reductase (GR) using nicotinamide adenine dinucleotide phosphate hydrogen (NADPH) as an electron donor. Dehydroascorbate may be reduced nonenzymatically or catalyzed by proteins with dehydroascorbate reductase (DHAR) activity.
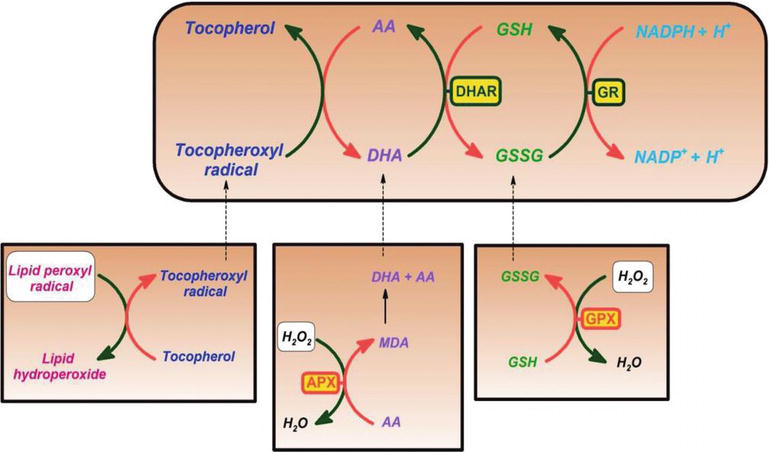
Figure 2.
xmlns:mml=”http://www.w3.org/1998/Math/MathML” xmlns:xlink=”http://www.w3.org/1999/xlink” xmlns:xsi=”http://www.w3.org/2001/XMLSchema-instanceAscorbate and redox cycling antioxidants. AA, ascorbate; DHA, dehydroascorbate; DHAR, semidehydroascorbate reductase; GSH, glutathione; GSSG, semi-glutathione reductase; GR, glutathione reductase; APX, ascorbate peroxidase; and GPX, glutathione peroxidase .
Glutathione-ascorbate cycle operates in the cytosol, mitochondria, plastids, and peroxisomes in plants . Det föreslås att glutation-askorbatcykeln spelar en nyckelroll för H2O2-avgiftning, på grund av de höga koncentrationerna av glutation, askorbat och NADPH i växtceller. Andra enzymer, såsom askorbat och glutationperoxidaser, som använder tioredoxiner eller glutaredoxiner som reducerande substrat, tar också roller i avlägsnandet av H2O2 i växter (Figur 2).
C-Vitamin bildar också semidehydroaskorbyl-radikalen, en relativt långlivad radikal, vid regenerering av E-vitamin från dess radikala form, liksom i rensande radikaler. Växt-och djurceller innehåller ett NADH-beroende semidehydroaskorbatreduktasenzym (EC 1.6.5.4), vilket reducerar radikalen tillbaka till C-vitamin genom att använda NADH som en källa till reduktionsmedel (Figur 2). Både enzymatiskt och nonenzymatiskt kan det irreversibelt sönderdelas till diketoglukonsyra eller det kan omvandlas till askorbat i en glutationberoende reaktion .
som en reducerande substans och en elektrondonator, under friradikalrensning, donerar C-vitamin högenergielektroner för att neutralisera fria radikaler, och det oxideras till dehydroaskorbinsyra. Dehydroaskorbinsyra kan omvandlas tillbaka till askorbinsyra för återanvändning eller kan metaboliseras, vilket ytterligare frigör fler elektroner. Även om C-vitamin absorberas från tarmen via en natriumberoende C-vitamintransportör, transporterar de flesta celler C-vitamin i en oxiderad form (dehydroaskorbinsyra) via glukostransportör 1. Dehydroaskorbinsyra reduceras för att generera askorbinsyra inuti cellen, vilket skyddar mitokondrier från friradikalinducerad oxidativ skada (figurerna 2 och 3). Mycket reaktiva fria radikaler (t. ex., RO–, RO2–, OH–,, NO2) reduceras med askorbat, och den nyligen genererade askorbylradikalen är dåligt reaktiv. Ascorbat kan också rensa icke-radikala reaktiva arter, härledda från peroxynitrit, såsom hypoklorsyra, ozon och nitreringsmedel. C-vitamin är en monosackaridoxidreducering (redox) katalysator som finns i både djur och växter. Antioxidanteffekten av C-vitamin beror på dess förmåga att donera elektroner från både andra och tredje kolet. Under primatutvecklingen har ett av enzymerna som behövs för att göra askorbinsyra gått förlorat genom mutation, människor måste få det från kosten ; de flesta djur kan syntetisera detta vitamin i sina kroppar och behöver inte det i sina dieter . C-Vitamin behövs vid omvandling av prokollagen till kollagen genom att oxidera prolinrester till hydroxiprolin. I andra celler upprätthålls den i sin reducerade form genom reaktion med glutation . Som visas i figurerna 2 och 3 är askorbinsyra en redoxkatalysator som kan reducera och därigenom neutralisera ROS såsom väteperoxid (H2O2) (figurerna 2 och 3).
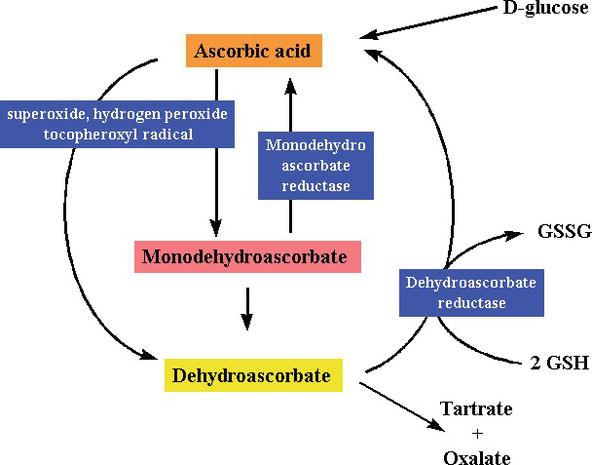
Figure 3.
xmlns:mml=”http://www.w3.org/1998/Math/MathML” xmlns:xlink=”http://www.w3.org/1999/xlink” xmlns:xsi=”http://www.w3.org/2001/XMLSchema-instanceSynthesis and degradation ofl-ascorbic acid in plant tissues .
Ascorbic acid has direct antioxidant effects, and also it is a substrate for the redox enzyme ascorbate peroxidase, that is particularly important in stress resistance in plants. Askorbinsyra är närvarande vid höga nivåer i alla delar av växter, särskilt i kloroplaster som når koncentrationer på 20 mM där . Dehydroascorbat (DHA) och askorbatfri radikal (AFR), som en mellanprodukt, askorbatfri radikal (AFR), som är reversibla, genereras enelektronoxidationer från askorbat (Figur 4). Enligt den allmänt antagna modellen för enzymatisk avlägsnande av ROS katalyserar SOD superoxidanjon till H2O2 och syre; sedan reduceras H2O2 till vatten och molekylärt syre av CAT. CAT turnover number is very high, but its affinity for H2O2 is relatively low, and consequently a certain amount of H2O2 remains in the cell.
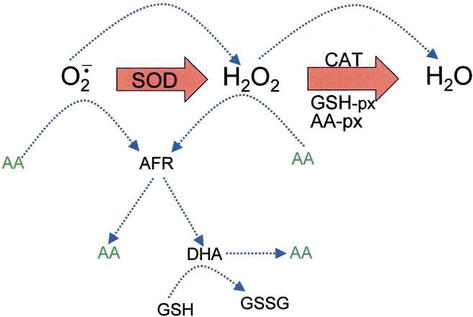
Figure 4.
xmlns:mml=”http://www.w3.org/1998/Math/MathML” xmlns:xlink=”http://www.w3.org/1999/xlink” xmlns:xsi=”http://www.w3.org/2001/XMLSchema-instanceThe role of AA in the detoxification of ROS. Blue dotted lines indicate nonenzymatic reactions.
H2O2 can react with superoxide anion formed in oxidative metabolism generating the highly reactive hydroxyl radical. GSH peroxidases (GSH-px) and AA peroxidases (AA-px) are capable of scavenging H2O2 due to their high affinity for H2O2. The cooperativity of SOD, CAT, and peroxidases ensures low amounts of superoxide anion and H2O2 and limiting the risk of hydroxyl radical formation (Figure 5).

Figure 5.
xmlns:mml=”http://www.w3.org/1998/Math/MathML” xmlns:xlink=”http://www.w3.org/1999/xlink” xmlns:xsi=”http://www.w3.org/2001/XMLSchema-instanceFoyer-Halliwell-Asada cycle .