Pyruvát kinázy M2 paliv více aspektů nádorových buněk: buněčný metabolismus, transkripční regulace extracelulární signalizace
Biochemické analýzy charactering enzymatickou aktivitu, která katalyzuje tvorbu laktátu z glukózy v buněčné lyzáty odhalil první intracelulární metabolické dráhy, glycolytic pathway. Počínaje od čištění frakcí, které obsahovaly glycolytic činnosti, počet pioneer vědci přispěli k identifikaci enzymů, které zahrnují v každém kroku v cestě . Tyto výsledky budují náš moderní koncept ve výměně aerobního a anaerobního dýchání a výroby energie za různých fyziologických a patologických okolností.
existence enzymu, který katalyzuje produkci ATP přenosem fosfátové skupiny z PEP na ADP v játrech byla poprvé zaznamenána v roce 1934 . Následná izolace enzymu, později známého jako pyruvát kináza (PK), prokázala rozdíly v tkáňové distribuci a katalytické kinetice, což naznačuje, že tento enzym může mít různé izoformy . Během let 1982 až 1984 byly různé PK geny klonovány z kvasinek, kuřat a potkanů . Funkční studie PKM2 byla zahájena identifikací kandidátského genu u myší na počátku 80 .let. Později, Noguchi et al. ukázalo se, že dvě izoformy PK (PKM1 a PKM2) jsou kódovány stejným genem PKM prostřednictvím alternativního sestřihu . U člověka jsou izoformy PKM také produkovány podobným spojovacím mechanismem zahrnutím exonu 9 a 10 do PKM1 a PKM2 mRNA odděleně .
několik zjištění upoutalo pozornost výzkumníka na potenciální roli PKM2 v tumorigenezi. Za prvé, PKM2 je embryonální izoforma, která je vysoce exprimována během vývoje zvířat. Jeho transkripce je oslabena v řadě dospělých tkání, zatímco je reaktivována v nádorech . Za druhé, studie relativní hojnosti PKM1 a PKM2 v normálních a nádorových tkáních prokázala přechod z izoformy PKM1 na izoformu PKM2 u různých rakovin, jako je hepatocelulární karcinom . Za třetí, změna mRNA sestřih z PKM1 na PKM2 je posílena tím, že c-Myc onkogenu naznačuje, rakovinné buňky se aktivně zapojily do tohoto přepínače, aby se vešly jejich požadavek v proliferaci a metabolismus . Začtvrté, modulace aktivity PKM2 aktivátory nebo inhibitory ovlivňují růst nádoru in vivo .
- první díl: PKM2 jako metabolický enzym v cytoplazmě
- druhá epizoda: PKM2 jako modulátor signalizace v cytoplazmě
- třetí díl: PKM2 jako transkripční regulátor v jádře
- čtvrtá epizoda: PKM2 jako extracelulární signalizační komunikátor
- The fifth episode: nezodpovězená nesrovnalost PKM2
- pokračující epizoda: je PKM2 biomarker rakoviny a cíl léku?
první díl: PKM2 jako metabolický enzym v cytoplazmě
Protože role PKM2 v metabolické kontrole glykolýzy v nádorových buňkách byla značně revidována , jen jsme shrnuli tři zásadní rozdíly mezi PKM1 – a PKM2-zprostředkovaná katalýza a buněčný metabolismus. Prvním rozdílem je interakce podjednotky. PKM1 i PKM2 jsou tetramerní proteiny tvořené čtyřmi identickými podjednotkami. Každá podjednotka (nebo monomer) obsahuje čtyři strukturní domény včetně a, B, C A N-terminální domény. Monomer nejprve dimerizuje společně a pak dva dimery interagují přes rozhraní dimer-dimer řízené doménou C monomeru za vzniku tetrameru. Protože PKM1 a PKM2 zahrnují různé exony ve svých mRNA, mění to kódované aminokyseliny v doméně C a mění stabilitu tetrameru. Za fyziologický stav, PKM1 constitutively organizuje jako tetramer, zatímco PKM2 může být existovala v tetramer nebo dimeru. Druhým rozdílem je všechnysterická regulace. V závislosti na intracelulárních koncentracích malých molekul a metabolitů může být aktivita PKM1 a PKM2 diferencovaně regulována. Jedním z nejznámějších alosterických regulátorů je fruktóza-1,6-bisfosfát (FBP). Tento glykolytický meziprodukt přímo váže PKM2 a zvyšuje afinitu PKM2 k PEP . Naopak, FBP významně neovlivňuje aktivitu PKM1. Kromě FBP bylo hlášeno, že jiné metabolity, aminokyseliny a malé molekuly ovlivňují aktivitu PKM2 (obr. 1). Koncentrace potřebná pro aktivaci nebo inhibici je však vysoká a modulační účinek je mírný. Zda tyto malé molekuly hrají důležitou roli při kontrole aktivity PKM za fyziologických okolností, je stále nejasné. PKM2 aktivita je také regulována post-translační modifikace, jako jsou fosforylace, acetylace a oxidace, které upřednostňují nízkou aktivitou dimerní PKM2 (Obr. 1). Třetím rozdílem je výroba energie a mezilehlé využití. Protože PKM1 konstitutivně existuje jako aktivní tetramer, hlavní biologickou funkcí této izoformy je generování ATP pro zásobování buněčné energie. Nicméně, PKM2, kromě toho k výrobě ATP, můžete přepnout na méně aktivní dimerní formě generovat několik glycolytic meziprodukty, které lze použít jako stavební bloky pro biosyntézu aminokyselin, lipidů a nukleotidů.
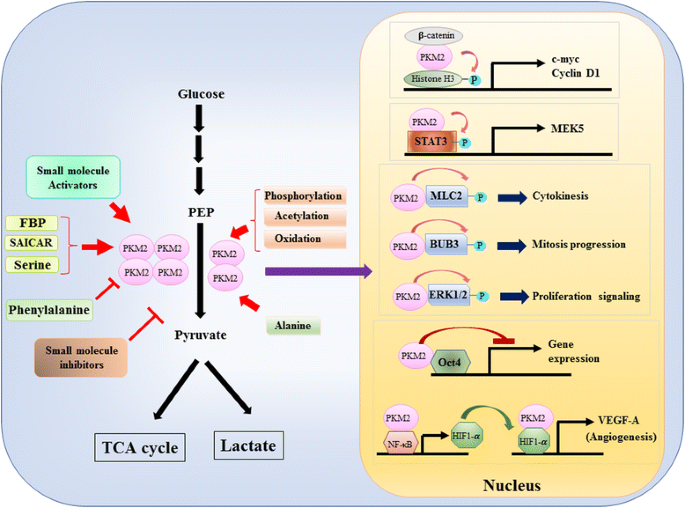
Modulace PKM2 aktivity podle fyziologických molekul a post-translační modifikace a potenciální roli PKM2 v transkripční regulace
druhá epizoda: PKM2 jako modulátor signalizace v cytoplazmě
kromě toho, aby fungoval jako glycolytic enzym, PKM2 se navrhuje zapojit do více buněčných procesů díky identifikaci interagujících proteinů v cytoplazmě. Například, PKM2 bylo prokázáno, že k interakci proteinu z několika tyrosin kináz včetně A-Raf, Break-point cluster region-Abelson (BCR-ABL) fúzní kinázy, fibroblastový růstový faktor receptor 1 (FGFR1) atd. . Ukázalo se, že tito vazební partneři modulují dimerní/tetramerní změnu PKM2 za účelem změny buněčného metabolismu. Je však možné, že PKM2 mohou vzájemně ovlivňují katalytické kinetiky, substrátové závazné a cytoplazmatické umístění těchto vazebných partnerů modulují signální transdukci. Zjištění, že PKM2 je fosfor-tyrosin vázající protein posiluje tuto možnost, protože mnoho intracelulární signální mediátory mohou vázat fosfor-tyrosin reziduí sestavit specifické proteinové komplexy pro přenos signálu . Seznam partnerů vázajících PKM2 k dnešnímu dni neustále roste. Zdůrazňujeme několik nových členů a diskutujeme zde o jejich biologických důsledcích. Mukherie a kol. prokázáno, že PKM2, že by mohla vázat s HuR, RNA vázající protein, který hraje důležitou roli v regulaci mRNA stabilitu a translační účinnost, podporovat progresi buněčného cyklu a proliferace gliom buněk . Zajímavé je, že další RNA vázající protein tristetraprolin, který může vázat několik mRNA prostřednictvím AU-bohaté prvek na 3′-untranslational regionu, (3′-UTR) bylo také zjištěno, že být PKM2 interakci partnera, a PKM2 indukované fosforylace a degradace tristetraprolin modulovat prsu růstu . Tyto dvě studie naznačují potenciální translační kontrolní funkci PKM2. Nedávno, Liang a kol. identifikoval antiapoptotický protein Bcl2 jako nového partnera PKM2 . Jsou prokázáno, že oxidační stres vyvolaný translokaci PKM2 do mitochondrií, kde je fosforylován a stabilizovaný Bcl2 tím, že zabraňuje jeho degradaci prostřednictvím ubikvitinace-závislé dráhy. Tyto údaje naznačují, že PKM2 pomáhá rakovinné buňky přizpůsobit oxidačním stresem vyvolané intracelulární metabolické změny nebo extracelulární urážet.
třetí díl: PKM2 jako transkripční regulátor v jádře
jaderné roli PKM2 v regulaci genové transkripce nebo epigenetické modifikace byla nejprve navrhl zjištění, že PKM2 vázán s Y333-phosphorlated β-catenin a β-catenin-PKM2 komplex byl přijat do nucleosomes, aby fosforylovat histonu H3 na threonin 11 (Obr. 1). Tato fosforylace následně zvýšila acetylaci histonu H3, což vedlo k upregulaci β-kateninových cílových genů. Další transkripční faktor přímo fosforylován PKM2 je signál snímače a aktivátor transkripce 3 (STAT3) . PKM2-zprostředkovaná fosforylace STAT3 na tyrosin 705 posílené aktivity STAT3, že upregulate výraz mitogen-aktivované protein kinázy kinázy 5 (MEK5). Vedle transkripčních faktorů, PKM2 bylo prokázáno, že fosforylují myosin light chain 2 (MLC2), BUB3 a extracelulární signálem regulované kinázy 1 a 2 (ERK1 a ERK2) . Zajímavé je, že PKM2 také působí způsobem nezávislým na fosforylaci a ovlivňuje expresi genů. Například, PKM2 bylo zjištěno, že vázat s Oct4, jeden z hlavní transkripční faktory, které kontrolují sebeobnovy kmenových buněk a potlačení Oct4-zprostředkované transkripce . PKM2 může také zvýšit nádorové angiogeneze interakcí s NF-kB a HIF-1α v jádře a aktivuje expresi HIF-1α cílových genů VEGF-A. v důsledku toho, zvýšená sekrece VEGF-A posiluje krevní cévy formace, která přispívá k růstu nádoru . I když tyto studie silně navrhl jaderné lokalizace a protein kinázy funkce PKM2 v různých fyziologických a patologických okolností však význam jaderné PKM2-zprostředkované genové exprese byl napadán studie pomocí PKM2 knockout buněk. Použitím-značených PEP a PKM2-null myších embryonálních fibroblastů, Hosios et al. ukázalo se, že fosforylace závislá na PEP není v buňkách běžnou událostí a reakce není katalyzována PKM2 . Rozpor těchto studií je v současné době nevyřešen a aktivita proteinkinázy PKM2 vyžaduje další potvrzení.
čtvrtá epizoda: PKM2 jako extracelulární signalizační komunikátor
přítomnost extracelulárního PKM2 otevírá novou cestu pro studium biologické funkce PKM2. Buschow a kol. poskytl první důkaz, že PKM2 může být detekován v B-buňce exosomy a byl identifikován jako MHC class II-associated protein . Dvě následující studie také ukázaly, že PKM2 existuje v exosomech uvolňovaných různými buňkami . V současné době, několik veřejných databází jako ExoCarta a EVpedia poskytovat komplexní informace pro součásti, včetně proteinů, lipidů, nukleových kyselin extracelulární váčky u různých druhů. Všechna data potvrzují, že PKM2 je obalový protein exosomů. Nedávné studie jasně prokázaly komunikační roli exosomů dodáním různých složek z hostitelských buněk do přijímajících buněk . Lze očekávat, že PKM2 může hrát roli v přeslech buněk.
vznikající důkazy tuto hypotézu skutečně podporují. Například nedávná studie prokázala, že PKM2 cirkulující v krvi může podporovat růst nádoru a angiogenezi zvýšením růstu, migrace a matrice adheze endotelových buněk . Další výzkum také ukázal, že PKM2 vylučovaný z buněk rakoviny tlustého střeva může působit autokrinní stimulací ke zvýšení migrace buněk aktivací dráhy PI3K / Akt a Wnt / β-Katenin . Kromě rakovinných buněk mohou neutrofily v místech poškození tkání uvolňovat PKM2, aby podpořily angiogenezi a hojení ran . Naše nedávná studie také prokázala, že rekombinantní PKM2 protein může indukovat fosforylaci a aktivaci receptoru epidermálního růstového faktoru (EGFR). Navíc jsme zjistili, že R339E mutant PKM2, který přednostně tvořil dimerní PKM2, aktivoval EGFR významněji než tetramerní PKM2. Keller et al. zjištěné 154 proteiny jako potenciální substráty pro PKM2 po ošetření Hela buněk s sukcinyl-5-aminoimidazole-4-karboxamid-1-ribóza-5′-fosfát (SAICAR), intracelulární metabolit, který by mohl stimulovat protein kinázy z PKM2 . Našli také EGFR jako substrát PKM2. Jejich výsledky se liší od našich dvěma způsoby. První, signální dráhy aktivované v naší studii jsou vyvolané extracelulární PKM2, zatímco molekulární cíle identifikované v jejich studii jsou potenciálními substráty intracelulárních PKM2. Za druhé, zvýšení aktivity ERK1 / 2 v naší studii je zahájeno aktivací EGFR, zatímco aktivace ERK1/2 v jejich studii je přímo stimulována komplexem SAICAR/PKM2. Jeden podobný jev pozorován v obou studiích je, že R339E mutant PKM2 aktivuje signální molekuly výrazněji než divoký typ PKM2 což naznačuje výraznou roli dimerní a tetrameric PKM2 v onkogenezi. Použitím receptorového tyrosinkinázového pole jsme zjistili, že extracelulární PKM2 aktivoval pouze omezené receptory růstového faktoru v buňkách rakoviny prsu (údaje nejsou zobrazeny). V současné době zůstává selektivita aktivace receptoru extracelulárním PKM2 neznámá. Kromě toho není jasné, proč je R339E mutant PKM2 účinnější při aktivaci EGFR. K zodpovězení těchto otázek je zapotřebí více experimentů.
Další elegantní otázku je třeba řešit, je, zda zdarma PKM2 a váček-balení PKM2 vykazují podobný účinek v podpoře tumorigenezi (Obr. 2). Volný extracelulární PKM2 nemohl proniknout do plazmatické membrány a mohl aktivovat intracelulární signalizaci pouze prostřednictvím buněčných povrchových proteinů, jako jsou receptory růstového faktoru. Naopak, váček-balení PKM2 mohl být endocytosed rakovinou a stromatu buněk, a PKM2 propuštěn z váčků může ovlivnit metabolismus a genovou expresi prostřednictvím intracelulární mechanismus. The PKM2-null cells or mice will be useful to elucidate whether these two extracellular forms of PKM2 could act synergistically or antagonistically in tumorigenesis.

The potential tumor-promoting effect of extracellular PKM2
The fifth episode: nezodpovězená nesrovnalost PKM2
kromě výše uvedených buněčných dat byla onkogenní role PKM2 také zpochybněna po generaci PKM2 knockoutových myší. Israelsen et al. vygeneroval podmíněný model knockout myši odstraněním exonu 10 specifického pro PKM2 . Překvapivě, deplece PKM2 urychlila, ale ne oslabila tvorbu nádorů způsobenou ztrátou genu Brca1 u myší. Tyto údaje ukázaly, že PKM2 není nutné pro proliferaci rakovinných buněk. Je zajímavé, že exprese PKM1 byla detekována pouze v neproliferujících nádorových buňkách, což naznačuje tumor supresivní úlohu PKM1 u rakoviny prsu. Kromě toho mají PKM2 knockout myši vysoký výskyt spontánního vývoje hepatocelulárního karcinomu po dlouhé latenci v důsledku nerovnováhy metabolismu . Tyto výsledky proti představě, že PKM2 hraje onkogenní roli in vivo.
pokračující epizoda: je PKM2 biomarker rakoviny a cíl léku?
i když výsledky geneticky inženýrství myš model nepodporují nádoru-podpora činnosti PKM2, zvýšená exprese PKM2 je všeobecně nalezen v lidských nádorových onemocnění a je spojena se špatným klinickým výsledkem (Tabulka 1) . Tento závěr podpořily také dvě nedávné metaanalytické studie. Wu a kol. analyzoval data z 2812 pacientů se solidními nádory trávicí soustavy získané od 16 kohortových studií a zjistili, že zvýšená exprese PKM2 je spojena se snížením celkové přežití u rakoviny žaludku, jícnu-spinocelulární karcinom, hepatocelulární karcinom, rakovina žlučových a rakovinu ústní dutiny . PKM2 však není prognostickým faktorem rakoviny pankreatu. Toto zjištění není v souladu s předchozími studiemi, které ukazují, že zvýšení PKM2 je indikátorem špatného přežití pacientů s rakovinou pankreatu . Další šetření zahrnovalo 4796 případů z 27 individuálních studií prokázalo, že upregulace PKM2 koreluje s horším celkovým přežitím, přežitím bez onemocnění a přežitím bez recidivy v souhrnných datech . Nicméně, rozvrstvené podle typu rakoviny, PKM2 nepředpovídá špatné přežití rakoviny pankreatu. Souhrnně se zdá, že PKM2 je odpovědným prognostickým markerem ve většině solidních nádorů.
naopak, použití PKM2 jako diagnostický faktor je kontroverzní. Proteomická analýza prokázala, že PKM2 je potenciálním diagnostickým markerem pro detekci rakoviny plic . Nedávná studie však naznačila, že PKM2 není dobrým diagnostickým markerem pro rakovinu plic kvůli nízké specificitě . Podobně je nepravděpodobné, že by samotný PKM2 byl užitečným markerem pro screening rakoviny tlustého střeva . Kombinace více markerů by však mohla zvýšit citlivost a specificitu pro diagnostiku rakoviny .
terapeutický potenciál PKM2 je zajímavou událostí v léčbě rakoviny. Z jedné strany se očekává, že inhibice PKM2 inhibuje glykolýzu, narušuje transkripci genů a potlačuje buněčnou proliferaci. Proto se zdá, že inhibitory PKM2 jsou dobrými kandidáty na vývoj protinádorových léčiv. Pomocí screeningu knihovny, Vander Heiden et al. identifikoval tři nové třídy inhibitorů PKM2 a ukázal, že nejúčinnější sloučenina inhibovala aktivitu PKM2 a indukovala smrt rakovinných buněk . Nedávno, Ning a kol. bylo zjištěno, že nové deriváty naftochinonu jsou silnými inhibitory PKM2 . Jedna účinná sloučenina 3 k potlačila proliferaci mnohočetných nádorových buněčných linií při submikromolárních koncentracích, zatímco vykazovala malý škodlivý účinek na normální buňky. Z druhé strany může aktivace PKM2 také inhibovat růst nádoru. Protože nízká aktivita PKM2 dimer je hlavní izoforma, která spouští glykolýza v cytoplazmě a genovou transkripci v jádře v nádorových buňkách, PKM2 aktivátory, které mohou podporovat tvorbu tetrameric PKM2 může přepnout glykolýza do mitochondrií dráhy a snížení jaderných vstupu k tlumení genové transkripce. Oba účinky zhoršují metabolickou poptávku a signalizaci podporující růst, která vede k regresi nádoru. Dvě průkopnické studie identifikovaly různé aktivátory PKM2 a charakterizovaly jejich specificitu in vitro . Následné studie prokázaly, že PKM2 aktivátory skutečně povýšen tetramer tvorbu a potlačil růst nádoru in vivo . Tyto výsledky naznačují, že aktivátory PKM2 by mohly být slibnými protinádorovými léky.
rezistence na chemoterapii je hlavní blokádou léčby rakoviny. Překonání inherentní chemorezistence rakovinných buněk je naléhavé pro klinický výzkum. Byly odhaleny role PKM2 v chemorezistenci rakovinných buněk a bylo prokázáno, že cílení PKM2 znovu senzibilizuje chemorezistentní rakovinné buňky. Nedávná studie ukázala, že CD44 interaguje s PKM2 a potlačuje aktivitu PKM2 zvýšením fosforylace PKM2 Tyr105. CD44 ablace indukovaná přechodu od aerobní glykolýzy na mitochondriální respirace a zvýšení reaktivních forem kyslíku (ROS) výroby, což má za následek zvýšení cisplatina citlivost v kolorektální nádorové buňky . Bylo prokázáno, že inhibice aktivity PKM2 potlačuje glykolýzu a překonává rezistenci na cisplatinu. Proto kombinace inhibitorů cisplatiny a PKM2 může být účinnou strategií pro chemoterapii (obr. 3). Bylo také hlášeno, že PKM2 se podílí na regulaci rezistence gemcitabinu v rakovinných buňkách pankreatu. Kim a spol. ukázal, že PKM2 podporuje gemcitabin odpor prostřednictvím negativní regulace p38-zprostředkovaná fosforylace p53, a proto snižuje transkripční aktivitu p53 a potlačuje expresi pro-apoptotických genů . Mimoděložní exprese R399E-PKM2, která přednostně tvoří dimerní PKM2, zvyšuje rezistenci rakovinných buněk pankreatu na gemcitabin. Tyto důkazy naznačují, že PKM2 přispívá k indukci rezistence na léčivo prostřednictvím nemetabolického mechanismu. Protože dimerní PKM2 působí hlavně jako transkripční regulátor v nádorových buňkách, PKM2 aktivátory, které podporují tvorbu tetrameric PKM2 mohou být účinné v potlačení non-metabolické funkce PKM2 a snižuje odolnost vůči gemcitabinem u karcinomu pankreatu buňky. Bylo také prokázáno, že jaderná PKM2 přispívá k rezistenci inhibitoru EGFR u kolorektálního karcinomu a rakoviny plic . Li et al. ukázal, že jaderné PKM2-zprostředkovaná fosforylace STAT3 snižuje citlivost kolorektálního karcinomu buněk gefitinib a narušení interakce jaderného PKM2 a STAT3 obnovena gefitinib citlivost v buňkách . Nedávno jiná studie prokázala, že PKM2 se translokuje do jádra a interaguje s poly-ADP ribózou (PAR) po stimulaci růstového faktoru. PAR-vazebná aktivita PKM2 je kritická pro jadernou retenci a genovou transkripci PKM2 a je důležitá pro podporu buněčné proliferace a růstu nádoru. Dospěli k závěru, že inhibice jaderné funkce PKM2 může překonat rezistenci rakovinných buněk mutovaných EGFR . Obě studie poukázaly na klíčovou roli jaderného PKM2 při zprostředkování rezistence na léky u rakovin a navrhly, že cílení jaderného PKM2 může být slibnou strategií k potlačení rezistence.

PKM2 přispívá k proliferace a lékové rezistence u nádorových buněk,