biokemisk analyse ved at karakterisere den fermatiske aktivitet, der katalyserer dannelsen af lactat fra glukose i cellelysater afslørede den første intracellulære metaboliske vej, den glykolytiske vej. Begyndende fra oprensningen af fraktioner, der indeholdt glycolytisk aktivitet, bidrog en række pionerforskere til identifikation af Fermes, der involverer i hvert trin i vejen . Disse resultater opbygger vores moderne koncept i udvekslingen af aerob og anaerob respiration og energiproduktion under forskellige fysiologiske og patologiske omstændigheder.i 1934 blev der først rapporteret om eksistensen af ATP, der katalyserede produktionen af ATP ved at overføre en fosfatgruppe fra PEP til ADP i leveren . Senere viste forskelle i vævsfordeling og katalytisk kinetik, hvilket tyder på, at dette kan have forskellige isoformer . I løbet af 1982 til 1984 blev forskellige PK-gener klonet fra gær, kylling og rotte . Den funktionelle undersøgelse af PKM2 blev initieret ved identifikation af et kandidatgen i mus i begyndelsen af 1980 ‘ erne . Senere, Noguchi et al. viste, at to isoformer af PK (PKM1 og PKM2) er kodet af det samme PKM-gen via alternativ splejsning . Hos mennesker produceres PKM-isoformer også via en lignende splejsningsmekanisme ved at inkludere ekson 9 og 10 i pkm1 og PKM2 mRNA separat .
flere fund fangede forskerens opmærksomhed på den potentielle rolle PKM2 i tumorigenese. For det første er PKM2 den embryonale isoform, der stærkt udtrykkes under dyreudvikling. Dens transkription dæmpes i et antal voksne væv, mens det genaktiveres i tumorer . For det andet viste undersøgelse af den relative overflod af PKM1 og PKM2 i normale og tumorvæv en skifte fra pkm1-isoformen til PKM2-isoformen i forskellige kræftformer som hepatocellulært carcinom . For det tredje forbedres ændringen af mRNA-splejsning fra PKM1 til PKM2 af C-Myc-onkogen, hvilket antyder, at kræftceller aktivt deltager i denne omskifter for at passe til deres krav til spredning og metabolisme . For det fjerde påvirker modulering af PKM2-aktivitet af aktivatorer eller hæmmere tumorvækst in vivo .
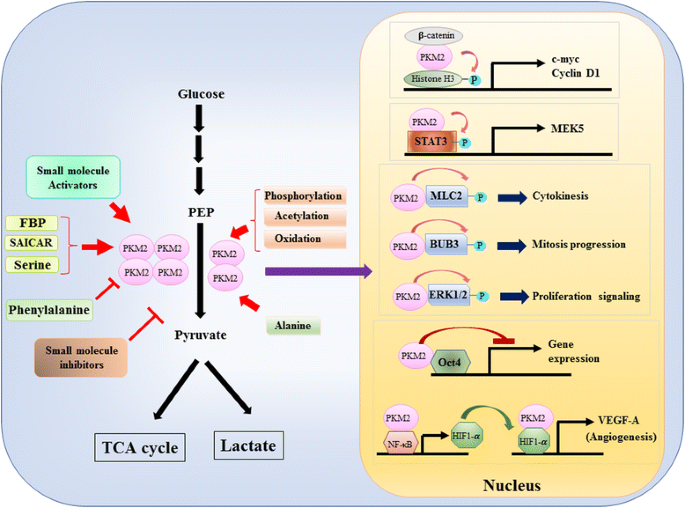
den første episode: Da pkm2 ‘ s rolle i metabolisk kontrol af glykolyse i kræftceller er blevet grundigt gennemgået , opsummerer vi kun tre afgørende forskelle mellem PKM1 – og PKM2-medieret katalyse og cellulær metabolisme her. Den første forskel er underenheds interaktion. Både PKM1 og PKM2 er tetramer proteiner dannet af fire identiske underenheder. Hver underenhed (eller monomer) indeholder fire strukturelle domæner, herunder A, B, C og N-terminal domæne. Monomeren dimeriserer først sammen, og derefter interagerer to dimerer via dimer-dimer-grænsefladen orkestreret af C-domænet af monomer for at danne en tetramer. Fordi PKM1 og PKM2 inkluderer forskellige eksoner i deres mRNA ‘ er, ændrer dette de kodede aminosyrer i C-domænet og ændrer tetramer-stabiliteten. Under fysiologisk tilstand organiserer PKM1 konstitutivt som en tetramer, mens PKM2 kan eksistere i tetramer eller dimer. Den anden forskel er allosterisk regulering. Afhængigt af de intracellulære koncentrationer af små molekyler og metabolitter kan aktiviteten af PKM1 og PKM2 reguleres differentielt. En af de mest kendte allosteriske regulatorer er fructose-1,6-bisphosphat (FBP). Dette glykolytiske mellemprodukt binder direkte PKM2 og øger affiniteten af PKM2 til PEP . Tværtimod påvirker FBP ikke signifikant pkm1-aktivitet. Ud over FBP er andre metabolitter, aminosyre og små molekyler rapporteret at påvirke PKM2-aktivitet (Fig. 1). Imidlertid er den koncentration, der kræves til aktivering eller inhibering, høj, og den modulerende effekt er beskeden. Hvorvidt disse små molekyler spiller en vigtig rolle i kontrollen af PKM-aktivitet under fysiologiske omstændigheder er stadig uklart. Pkm2-aktivitet reguleres også ved posttranslationel modifikation, såsom phosphorylering, acetylering og iltning, som favoriserer den lave aktivitet af dimerisk PKM2 (Fig. 1). Den tredje forskel er energiproduktion og mellemudnyttelse. Da PKM1 eksisterer konstitutivt som den aktive tetramer, er den vigtigste biologiske funktion af denne isoform dannelsen af ATP til at levere cellulær energi. Imidlertid kan PKM2 ud over at producere ATP skifte til den mindre aktive dimeriske form for at generere flere glycolytiske mellemprodukter, der kan bruges som byggesten til biosyntese af aminosyrer, lipider og nukleotider.
Fig. 1
modulering af PKM2 aktivitet ved fysiologiske molekyler og post-translationel modifikation og den potentielle rolle PKM2 i transkriptionel regulering
den anden episode: pkm2 som signalmodulator i cytoplasmaet
ud over at fungere som et glykolytisk ferment foreslås pkm2 at involvere i flere cellulære processer på grund af identifikationen af interagerende proteiner i cytoplasmaet. For eksempel blev PKM2 vist at være et interagerende protein af flere tyrosinkinaser inklusive A-Raf, Brudpunkt klyngeregion-Abelson (BCR-ABL) fusionskinase, fibroblastvækstfaktorreceptor 1 (FGFR1) etc. . Disse bindingspartnere har vist sig at modulere den dimeriske/tetrameriske ændring af PKM2 for at ændre cellemetabolismen. Det er imidlertid muligt, at PKM2 gensidigt kan påvirke den katalytiske kinetik, substratbinding og cytoplasmatisk placering af disse bindingspartnere for at modulere signaltransduktion. Konstateringen af, at PKM2 er et fosfor-tyrosinbindende protein, styrker denne mulighed, fordi mange intracellulære signalmediatorer kan binde til fosfor-tyrosinrest for at samle specifikke proteinkomplekser til signaloverførsel . Til dato vokser listen over pkm2-bindende partnere kontinuerligt. Vi fremhæver flere nye medlemmer og diskuterer deres biologiske implikation her. Mukheriee et al. demonstreret, at PKM2 kunne binde med HuR, et RNA-bindende protein, der spiller en vigtig rolle i kontrollen af mRNA-stabilitet og translationel effektivitet for at fremme cellecyklusprogression og proliferation af gliomceller . Interessant nok blev et andet RNA-bindende protein tristetraprolin, som kunne binde et antal mRNA via det AU-rige element ved 3′-ikke-Translational region (3′ – UTR) også fundet at være en pkm2-interagerende partner, og PKM2 induceret phosphorylering og nedbrydning af tristetraprolin for at modulere brystkræftvækst . Disse to undersøgelser indebærer en potentiel translationel kontrolfunktion af PKM2. For nylig, Liang et al. identificeret det anti-apoptotiske protein Bcl2 som en ny pkm2-partner . De viste, at oksidativ stress inducerede translokationen af PKM2 til mitokondrier, hvor den phosphorylerede og stabiliserede Bcl2 ved at forhindre dens nedbrydning via allestedsnærværende afhængig vej. Disse data antydede, at PKM2 hjælper kræftceller med at tilpasse sig oksidativ stress fremkaldt af intracellulær metabolisk ændring eller ekstracellulær fornærmelse.
den tredje episode: PKM2 som en transkriptionsregulator i kernen
en nuklear rolle af PKM2 i reguleringen af gentranskription eller epigenetisk modifikation blev for det første foreslået ved konstateringen af, at PKM2 bundet med Y333-phosphorlateret Purpur-catenin, og at komplekset med Purpur-catenin-PKM2 blev rekrutteret til nukleosomerne for at phosphorylere histon H3 ved threonin 11 (Fig. 1). Denne phosphorylering øgede efterfølgende histon H3-acetylering, der førte til opregulering af purpur-catenin-målgener. En anden transkriptionsfaktor direkte phosphoryleret af PKM2 er signaltransducer og aktivator af transkription 3 (STAT3) . PKM2-medieret phosphorylering af STAT3 ved tyrosin 705 forbedrede STAT3-aktiviteten til opregulering af ekspressionen af mitogenaktiveret proteinkinase kinase 5 (MEK5). Ved siden af transkriptionsfaktorer har PKM2 vist sig at phosphorylere myosin let kæde 2 (MLC2), BUB3 og ekstracellulær signalreguleret kinase 1 og 2 (ERK1 og ERK2) . Interessant nok virker PKM2 også via phosphoryleringsuafhængig måde for at påvirke genekspression. For eksempel har PKM2 vist sig at binde med Oct4, en af mastertranskriptionsfaktorerne, der styrer selvfornyelse af stamceller og hæmmer Oct4-medieret transkription . PKM2 kan også forbedre tumorangiogenese ved at interagere med NF-kB og HIF-1-kur i kernen og aktivere ekspressionen af HIF-1-target-genet VEGF-A. følgelig øger øget sekretion af VEGF-A blodkardannelse, som bidrager til tumorvækst . Selvom disse undersøgelser stærkt antydede nuklear lokalisering og proteinkinasefunktion af PKM2 under forskellige fysiologiske og patologiske omstændigheder, er vigtigheden af nuklear PKM2-medieret genekspression imidlertid blevet udfordret af undersøgelser, der bruger pkm2 knockout-celler. Ved at bruge-mærket PEP og PKM2-null mus embryonale fibroblaster, Hosios et al. viste, at PEP-afhængig phosphorylering ikke er en almindelig begivenhed i celler, og reaktionen katalyseres ikke af PKM2 . Uoverensstemmelsen mellem disse undersøgelser er i øjeblikket uløst, og proteinkinaseaktiviteten af PKM2 har brug for yderligere bekræftelse.
den fjerde episode: PKM2 som en ekstracellulær signalkommunikator
tilstedeværelsen af ekstracellulær PKM2 åbner en ny vej til undersøgelse af pkm2 biologisk funktion. Buschu et al. fremlagde det første bevis for, at PKM2 kunne påvises i B-celleeksosomer og blev identificeret som et MHC klasse II-associeret protein . To efterfølgende undersøgelser viste også, at PKM2 eksisterede i eksosomer frigivet af forskellige celler . I øjeblikket giver flere offentlige databaser som f.eksocarta og EVpedia omfattende information om komponenterne, herunder proteiner, lipider, nukleinsyrer af ekstracellulære vesikler i forskellige arter. Alle data bekræfter, at PKM2 er et pakkeprotein af eksosomer. Nylige undersøgelser har tydeligt vist en kommunikativ rolle af eksosomer ved at levere forskellige komponenter fra værtsceller til modtagerceller . Det kan forventes, at PKM2 kan spille en rolle i Celle-Celle krydstale.
nye beviser understøtter faktisk denne hypotese. For eksempel viste en nylig undersøgelse, at blod cirkulerende PKM2 kan fremme tumorvækst og angiogenese ved at øge væksten, migrationen og matricen adhæsion af endotelceller . En anden undersøgelse viste også, at PKM2, der udskilles fra tyktarmscancerceller, muligvis virker via en autokrin stimulering for at forbedre cellemigration ved at aktivere PI3K/Akt og vnt/LARP-catenin-veje . Ud over kræftceller kunne neutrofiler på vævsskadestederne frigive PKM2 for at fremme angiogenese og sårheling . Vores nylige undersøgelse viste også, at rekombinant pkm2-protein kunne inducere phosphorylering og aktivering af epidermal vækstfaktorreceptor (EGFR) . Desuden fandt vi, at R339E mutant PKM2, som fortrinsvis dannede dimerisk PKM2, aktiverede EGFR mere signifikant end den tetrameriske PKM2. Keller et al. identificerede 154 proteiner som potentielle substrater for PKM2 efter behandling af Hela-celler med succinyl-5-aminoimidasol-4-carbosamid-1-ribose-5′ – phosphat (SAICAR), en intracellulær metabolit, der kunne stimulere proteinkinaseaktiviteten af PKM2 . De fandt også EGFR som et pkm2-substrat. Deres resultater er forskellige fra vores på to måder. For det første fremkaldes de signalveje, der er aktiveret i vores undersøgelse, af ekstracellulær PKM2, mens de molekylære mål, der er identificeret i deres undersøgelse, er potentielle substrater for intracellulær PKM2. For det andet er stigningen i ERK1/2-aktivitet i vores undersøgelse initieret af EGFR-aktivering, mens ERK1/2-aktivering i deres undersøgelse stimuleres direkte af SAICAR/PKM2-komplekset. Et lignende fænomen observeret i begge undersøgelser er, at R339E mutant PKM2 aktiverer signalmolekyler mere signifikant end den vilde type PKM2, hvilket antyder den særskilte rolle af dimer og tetramer PKM2 i onkogenese. Ved at bruge receptortyrosinkinase array fandt vi, at ekstracellulær PKM2 kun aktiverede begrænsede vækstfaktorreceptorer i brystkræftceller (data ikke vist). I øjeblikket forbliver selektiviteten af receptoraktivering ved ekstracellulær PKM2 ukendt. Derudover er det heller ikke klart, hvorfor R339E mutant PKM2 er mere potent i aktiveringen af EGFR. Flere eksperimenter er nødvendige for at besvare disse spørgsmål.
et andet elegant spørgsmål, der skal behandles, er, om fri PKM2 og vesikelemballage PKM2 udviser lignende virkning til fremme af tumorigenese (Fig. 2). Fri ekstracellulær PKM2 kunne ikke trænge ind i plasmamembranen og kunne kun aktivere intracellulær signalering via celleoverfladeproteiner som vækstfaktorreceptorer. Omvendt kunne vesikelemballage PKM2 endocytoseres af kræft og stroma celler, og PKM2 frigivet fra vesikler kunne påvirke metabolisme og genekspression via intracellulær mekanisme. The PKM2-null cells or mice will be useful to elucidate whether these two extracellular forms of PKM2 could act synergistically or antagonistically in tumorigenesis.
Fig. 2
The potential tumor-promoting effect of extracellular PKM2
The fifth episode: ubesvaret uoverensstemmelse mellem PKM2
ud over de cellebaserede data, der er diskuteret ovenfor, er pkm2 ‘ s onkogene rolle også blevet udfordret efter genereringen af pkm2 knockout-mus. Israelsen et al. genereret en betinget knockout-musemodel ved at slette den PKM2-specifikke ekson 10 . Overraskende accelererede udtømning af PKM2, men ikke svækket tumordannelse drevet af tab af Brca1-gen hos mus. Disse data viste, at PKM2 ikke er nødvendig for spredning af kræftceller. Interessant nok blev pkm1-ekspression kun påvist i ikke-prolifererende tumorceller, hvilket tyder på en tumorundertrykkende rolle af PKM1 i brystkræft. Derudover har pkm2 knockout-mus en høj forekomst til at udvikle hepatocellulært carcinom spontant efter en lang latenstid på grund af ubalancen i stofskiftet . Disse resultater mod forestillingen om, at PKM2 spiller en onkogen rolle in vivo.
den fortsatte episode: er PKM2 en kræftbiomarkør og lægemiddelmål?
selvom resultaterne af genetisk ingeniør musemodel ikke understøtter den tumorfremmende aktivitet af PKM2, findes overekspression af PKM2 universelt i humane kræftformer og er forbundet med dårligt klinisk resultat (tabel 1) . To nylige metaanalysestudier understøttede også denne konklusion. Et al. analyserede dataene fra 2812 patienter med faste tumorer i fordøjelsessystemet opnået fra 16 kohortestudier og fandt ud af, at overekspression af PKM2 er forbundet med reduceret samlet overlevelse i gastrisk kræft, esophageal pladecellecarcinom, hepatocellulært carcinom, galdecancer og oral kræft . PKM2 er imidlertid ikke en prognostisk faktor for kræft i bugspytkirtlen. Dette fund er ikke i overensstemmelse med tidligere undersøgelser, der viser, at stigning i PKM2 er en indikator for dårlig overlevelse hos patienter med kræft i bugspytkirtlen . En anden undersøgelse omfattede 4796 tilfælde fra 27 individuelle undersøgelser viste, at pkm2 opregulering er korreleret med dårligere samlet overlevelse, sygdomsfri overlevelse og gentagelsesfri overlevelse i poolede data . Stratificeret efter kræfttype forudsiger PKM2 imidlertid ikke en dårlig overlevelse af kræft i bugspytkirtlen. Samlet set synes PKM2 at være en ansvarlig prognostisk markør i de fleste solide tumorer.
tabel 1 prognostisk betydning af PKM2 i humane kræftformer
tværtimod er brugen af PKM2 som en diagnostisk faktor kontroversiel. En proteomisk analyse viste, at PKM2 er en potentiel diagnostisk markør til påvisning af lungekræft . En nylig undersøgelse antydede imidlertid, at PKM2 ikke er en god diagnostisk markør for lungekræft på grund af lav specificitet . Tilsvarende er PKM2 alene sandsynligvis ikke en nyttig markør til screening af tyktarmskræft . Imidlertid kan kombination af flere markører øge følsomheden og specificiteten for kræftdiagnose .
det terapeutiske potentiale af PKM2 er en spændende begivenhed i kræftbehandling. Fra den ene side forventes inhibering af PKM2 at hæmme glykolyse, forringe gentranskription og undertrykke cellulær proliferation. Derfor synes PKM2-hæmmere at være gode kandidater til udvikling af lægemidler mod kræft. Ved at bruge biblioteksscreening, Vander Heiden et al. identificerede tre nye klasser af PKM2-hæmmere og viste, at den mest effektive forbindelse hæmmede pkm2-aktivitet og induceret død af kræftceller . For nylig, Ning et al. fundet, at nye naphthokinonderivater er potente PKM2-hæmmere . En effektiv forbindelse 3 k undertrykte proliferationen af flere kræftcellelinjer ved submikromolære koncentrationer, mens den viste ringe skadelig virkning på normale celler. Fra den anden side kan aktivering af PKM2 også hæmme tumorvækst. Fordi PKM2-dimer med lav aktivitet er den største isoform, der udløser glykolyse i cytoplasmaet og gentranskription i kernen i kræftceller, kan PKM2-aktivatorer, der kan fremme dannelsen af tetramerisk PKM2, skifte glykolyse til mitokondriervej og reducere nuklear adgang for at dæmpe gentranskription. Begge effekter forringer metabolisk efterspørgsel og vækststøttende signalering, der fører til tumorregression. To pionerundersøgelser identificerede forskellige PKM2-aktivatorer og karakteriserede deres specificitet in vitro . En efterfølgende undersøgelse viste, at PKM2-aktivatorer faktisk fremmede dannelse af tetramer og undertrykte tumorvækst in vivo . Disse resultater antydede, at PKM2-aktivatorer kunne være lovende lægemidler mod kræft.
resistens over for kemoterapi er en stor blokering for kræftbehandling. At overvinde den iboende kemoresistens af kræftceller er presserende for klinisk forskning. Pkm2 ‘ s roller i kemoresistens af kræftceller er blevet afsløret, og målretning af PKM2 har vist sig at gensensibilisere kemoresistente kræftceller. En nylig undersøgelse viste, at CD44 interagerer med PKM2 og undertrykker PKM2-aktivitet via stigende Tyr105-phosphorylering af PKM2. CD44-ablation inducerede skiftet fra aerob glykolyse til mitokondriel respiration og stigende produktion af reaktive iltarter (ROS), hvilket resulterede i forbedring af cisplatinfølsomhed i kolorektale kræftceller . Inhibering af pkm2-aktivitet blev påvist at undertrykke glykolyse og overvinde cisplatinresistens. Derfor kan kombinationen af cisplatin og PKM2-hæmmere være en effektiv strategi for kemoterapi (Fig. 3). PKM2 er også rapporteret at deltage i reguleringen af gemcitabinresistens i bugspytkirtelkræftceller. Kim et al. viste, at PKM2 fremmer gemcitabinresistens gennem negativ regulering af p38-medieret p53-phosphorylering, hvilket reducerer transkriptionsaktiviteten af p53 og undertrykker ekspressionen af pro-apoptotiske gener . Desuden øger ektopisk ekspression af R399E-PKM2, som fortrinsvis danner dimerisk PKM2, resistensen af pancreascancerceller til gemcitabin. Disse beviser antyder, at PKM2 bidrager til induktion af lægemiddelresistens via en ikke-metabolisk mekanisme. Fordi dimerisk PKM2 fungerer hovedsageligt som en transkriptionsregulator i kræftceller, kan PKM2-aktivatorer, der fremmer dannelsen af tetramerisk PKM2, være effektive til at undertrykke ikke-metabolisk funktion af PKM2 og reducere resistens over for gemcitabin i bugspytkirtelkræftceller. Nuklear PKM2 har også vist sig at bidrage til modstanden af EGFR-hæmmer i kolorektal cancer og lungekræft . Li et al. viste, at nuklear PKM2-medieret STAT3 phosphorylering reducerer følsomheden af kolorektal cancerceller til gefitinib og forstyrrelse af interaktionen mellem nuklear PKM2 og STAT3 gendannet Gefitinib følsomhed i cellerne . For nylig viste en anden undersøgelse, at PKM2 translokeres i kernen og interagerer med poly-ADP ribose (PAR) ved vækstfaktorstimulering. Den PAR-bindende aktivitet af PKM2 er kritisk for nuklear retention og gentranskription af PKM2 og er vigtig for fremme af celleproliferation og tumorvækst. De konkluderede, at hæmning af pkm2 nuklear funktion kan overvinde modstanden af EGFR-muterede kræftceller . Begge undersøgelser påpegede en afgørende rolle for nuklear PKM2 i formidling af lægemiddelresistens i kræft og foreslået at målrette nuklear PKM2 kan være en lovende strategi for at tilsidesætte modstanden.
Fig. 3
PKM2 bidrager til proliferation og lægemiddelresistens i kræftceller