Pyruvatkinase M2 fördert mehrere Aspekte von Krebszellen: vom Zellstoffwechsel, Transkriptionsregulation zur extrazellulären Signalgebung
Die biochemische Analyse durch Charakterisierung der enzymatischen Aktivität, die die Bildung von Laktat aus Glucose in Zelllysaten katalysiert, ergab den ersten intrazellulären Stoffwechselweg, den glykolytischen Weg. Ausgehend von der Reinigung von Fraktionen, die glykolytische Aktivität enthielten, trugen eine Reihe von Pionierforschern zur Identifizierung von Enzymen bei, die an jedem Schritt des Weges beteiligt sind . Diese Ergebnisse bilden unser modernes Konzept im Austausch der aeroben und anaeroben Atmung und Energieproduktion unter verschiedenen physiologischen und pathologischen Umständen.Die Existenz eines Enzyms, das die Produktion von ATP katalysierte, indem es eine Phosphatgruppe von PEP zu ADP in der Leber übertrug, wurde erstmals 1934 berichtet . Die anschließende Isolierung des Enzyms, später bekannt als Pyruvatkinase (PK), zeigte Unterschiede in der Gewebeverteilung und der katalytischen Kinetik, was darauf hindeutet, dass dieses Enzym unterschiedliche Isoformen aufweisen kann . In den Jahren 1982 bis 1984 wurden verschiedene PK-Gene aus Hefe, Huhn und Ratte kloniert . Die funktionelle Studie von PKM2 wurde durch die Identifizierung eines Kandidatengens in der Maus in den frühen 1980er Jahren initiiert . Später Noguchi et al. zeigte, dass zwei Isoformen von PK (PKM1 und PKM2) durch das gleiche PKM-Gen über alternatives Spleißen kodiert werden. Beim Menschen werden PKM-Isoformen auch über einen ähnlichen Spleißmechanismus hergestellt, indem Exon 9 und 10 separat in PKM1- und PKM2-mRNA eingeschlossen werden .
Mehrere Ergebnisse erregten die Aufmerksamkeit der Forscher auf die mögliche Rolle von PKM2 bei der Tumorentstehung. Erstens ist PKM2 die embryonale Isoform, die während der Tierentwicklung exprimiert wird. Seine Transkription wird in einer Reihe von adulten Geweben abgeschwächt, während es in Tumoren reaktiviert wird . Zweitens zeigte die Untersuchung der relativen Häufigkeit von PKM1 und PKM2 in Normal- und Tumorgeweben einen Wechsel von der PKM1-Isoform zur PKM2-Isoform bei verschiedenen Krebsarten wie dem hepatozellulären Karzinom . Drittens wird der Wechsel des mRNA-Spleißens von PKM1 zu PKM2 durch das c-Myc-Onkogen verstärkt, was darauf hindeutet, dass Krebszellen aktiv an diesem Wechsel teilnehmen, um ihren Anforderungen an Proliferation und Metabolismus gerecht zu werden . Viertens beeinflusst die Modulation der PKM2-Aktivität durch Aktivatoren oder Inhibitoren das Tumorwachstum in vivo .
- Die erste Episode: PKM2 als metabolisches Enzym im Zytoplasma
- Die zweite Episode: PKM2 als Signalweg modulator im Zytoplasma
- Die dritte Episode: PKM2 als Transkriptionsregulator im Kern
- Die vierte Episode: PKM2 als extrazellulärer Signalkommunikator
- The fifth episode: unbeantwortete Diskrepanz von PKM2
- Die anhaltende Episode: Ist PKM2 ein Krebsbiomarker und ein Wirkstoffziel?
Die erste Episode: PKM2 als metabolisches Enzym im Zytoplasma
Da die Rolle von PKM2 bei der metabolischen Kontrolle der Glykolyse in Krebszellen ausführlich untersucht wurde, fassen wir hier nur drei entscheidende Unterschiede zwischen PKM1- und PKM2-vermittelter Katalyse und zellulärem Metabolismus zusammen. Der erste Unterschied ist die Untereinheitsinteraktion. Sowohl PKM1 als auch PKM2 sind tetramere Proteine, die aus vier identischen Untereinheiten bestehen. Jede Untereinheit (oder Monomer) enthält vier strukturelle Domänen, einschließlich A, B, C und N-terminale Domäne. Das Monomer dimerisiert zuerst zusammen und dann interagieren zwei Dimere über die Dimer-Dimer-Grenzfläche, die von der C-Domäne des Monomers orchestriert wird, um ein Tetramer zu bilden. Da PKM1 und PKM2 unterschiedliche Exons in ihren mRNAs enthalten, verändert dies die codierten Aminosäuren in der C-Domäne und verändert die Tetramerstabilität. Unter physiologischen Bedingungen organisiert sich PKM1 konstitutiv als Tetramer, während PKM2 in Tetramer oder Dimer vorhanden sein kann. Der zweite Unterschied ist die allosterische Regulation. Abhängig von den intrazellulären Konzentrationen von kleinen Molekülen und Metaboliten kann die Aktivität von PKM1 und PKM2 unterschiedlich reguliert werden. Einer der bekanntesten allosterischen Regulatoren ist Fructose-1,6-bisphosphat (FBP). Dieses glykolytische Zwischenprodukt bindet direkt PKM2 und erhöht die Affinität von PKM2 für PEP . Im Gegenteil, FBP beeinflusst die PKM1-Aktivität nicht signifikant. Zusätzlich zu FBP wurde berichtet, dass andere Metaboliten, Aminosäuren und kleine Moleküle die PKM2-Aktivität beeinflussen (Abb. 1). Die zur Aktivierung oder Hemmung erforderliche Konzentration ist jedoch hoch und die modulatorische Wirkung gering. Ob diese kleinen Moleküle unter physiologischen Umständen eine wichtige Rolle bei der Kontrolle der PKM-Aktivität spielen, ist noch unklar. Die PKM2-Aktivität wird auch durch posttranslationale Modifikationen wie Phosphorylierung, Acetylierung und Oxidation reguliert, die die geringe Aktivität von dimerem PKM2 begünstigen (Abb. 1). Der dritte Unterschied besteht in der Energieerzeugung und der Zwischennutzung. Da PKM1 konstitutiv als aktives Tetramer existiert, ist die biologische Hauptfunktion dieser Isoform die Erzeugung von ATP zur Versorgung der zellulären Energie. PKM2 kann jedoch nicht nur ATP produzieren, sondern auch in die weniger aktive dimere Form wechseln, um mehrere glykolytische Zwischenprodukte zu erzeugen, die als Bausteine für die Biosynthese von Aminosäuren, Lipiden und Nukleotiden verwendet werden können.
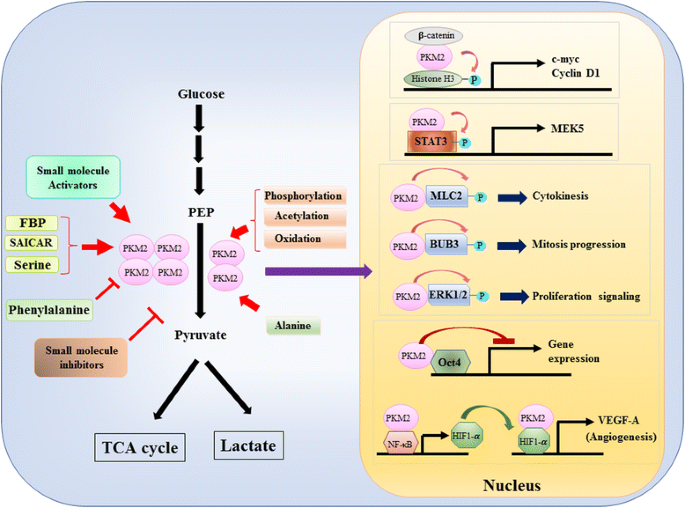
Modulation der PKM2-Aktivität durch physiologische Moleküle und posttranslationale Modifikation und die mögliche Rolle von PKM2 bei der Transkriptionsregulation
Die zweite Episode: PKM2 als Signalweg modulator im Zytoplasma
Zusätzlich zur Funktion als glykolytisches Enzym wird vorgeschlagen, dass PKM2 aufgrund der Identifizierung interagierender Proteine im Zytoplasma an mehr zellulären Prozessen beteiligt ist. Zum Beispiel wurde gezeigt, dass PKM2 ein interagierendes Protein mehrerer Tyrosinkinasen ist, einschließlich A-Raf, Break Point Cluster Region-Abl (BCR-ABL) Fusionskinase, Fibroblastenwachstumsfaktorrezeptor 1 (FGFR1) usw. . Es wurde gezeigt, dass diese Bindungspartner die dimere / tetramere Veränderung von PKM2 modulieren, um den Zellstoffwechsel zu verändern. Es ist jedoch möglich, dass PKM2 die katalytische Kinetik, Substratbindung und zytoplasmatische Lage dieser Bindungspartner wechselseitig beeinflusst, um die Signaltransduktion zu modulieren. Der Befund, dass PKM2 ein Phosphor-Tyrosin-Bindungsprotein ist, verstärkt diese Möglichkeit, da viele intrazelluläre Signalmediatoren an Phosphor-Tyrosin-Rückstände binden können, um spezifische Proteinkomplexe für die Signalübertragung zusammenzubauen . Bis heute wächst die Liste der PKM2-Bindungspartner kontinuierlich. Wir heben einige neue Mitglieder hervor und diskutieren hier ihre biologischen Implikationen. Mukheriee et al. gezeigt, dass PKM2 an HuR binden kann, ein RNA-Bindungsprotein, das eine wichtige Rolle bei der Kontrolle der mRNA-Stabilität und der Translationseffizienz spielt, um das Fortschreiten des Zellzyklus und die Proliferation von Gliomzellen zu fördern . Interessanterweise wurde ein weiteres RNA-bindendes Protein Tristetraprolin, das eine Reihe von mRNA über das AU-reiche Element an der 3′-untranslationalen Region (3′-UTR) binden konnte, ebenfalls als PKM2-Interaktionspartner gefunden, und PKM2 induzierte Phosphorylierung und Abbau von Tristetraprolin, um das Brustkrebswachstum zu modulieren . Diese beiden Studien implizieren eine mögliche translationale Kontrollfunktion von PKM2. Kürzlich haben Liang et al. identifizierte das anti-apoptotische Protein Bcl2 als neuen PKM2-Partner . Sie zeigten, dass oxidativer Stress die Translokation von PKM2 in Mitochondrien induzierte, wo es Bcl2 phosphorylierte und stabilisierte, indem es seinen Abbau über den ubiquitinierungsabhängigen Weg verhinderte. Diese Daten deuten darauf hin, dass PKM2 Krebszellen hilft, oxidativen Stress anzupassen, der durch intrazelluläre metabolische Veränderungen oder extrazelluläre Insult hervorgerufen wird.
Die dritte Episode: PKM2 als Transkriptionsregulator im Kern
Eine nukleare Rolle von PKM2 bei der Regulation der Gentranskription oder epigenetischen Modifikation wurde zunächst durch den Befund nahe gelegt, dass PKM2 an Y333-phosphorliertes β-Catenin gebunden war und der β-Catenin-PKM2-Komplex an die Nukleosomen rekrutiert wurde, um Histon H3 an Threonin 11 zu phosphorylieren (Abb. 1). Diese Phosphorylierung erhöhte anschließend die Histon-H3-Acetylierung, was zu einer Hochregulation von β-Catenin-Zielgenen führte. Ein weiterer Transkriptionsfaktor, der direkt durch PKM2 phosphoryliert wird, ist Signalwandler und Aktivator der Transkription 3 (STAT3) . Die PKM2-vermittelte Phosphorylierung von STAT3 an Tyrosin 705 erhöhte die STAT3-Aktivität, um die Expression der mitogenaktivierten Proteinkinase Kinase 5 (MEK5) zu regulieren. Neben Transkriptionsfaktoren phosphoryliert PKM2 nachweislich Myosin Light Chain 2 (MLC2), BUB3 und die extrazelluläre signalregulierte Kinase 1 und 2 (ERK1 und ERK2) . Interessanterweise wirkt PKM2 auch phosphorylierungsunabhängig, um die Genexpression zu beeinflussen. Beispielsweise wurde festgestellt, dass PKM2 an Oct4 bindet, einen der Haupttranskriptionsfaktoren, die die Selbsterneuerung von Stammzellen steuern, und die Oct4-vermittelte Transkription hemmt . PKM2 kann auch die Tumorangiogenese verbessern, indem es mit NF-kB und HIF-1α im Zellkern interagiert und die Expression des HIF-1α-Zielgens VEGF-A aktiviert. Obwohl diese Studien die nukleare Lokalisation und Proteinkinase-Funktion von PKM2 unter verschiedenen physiologischen und pathologischen Umständen stark nahelegten, wurde die Bedeutung der nuklearen PKM2-vermittelten Genexpression durch Studien mit PKM2-Knockout-Zellen in Frage gestellt. Durch die Verwendung von -markierten PEP- und PKM2-Null-mausembryonalen Fibroblasten, Hosios et al. zeigte, dass PEP-abhängige Phosphorylierung kein häufiges Ereignis in Zellen ist und die Reaktion nicht durch PKM2 katalysiert wird . Die Diskrepanz dieser Studien ist derzeit ungelöst und die Proteinkinaseaktivität von PKM2 muss weiter bestätigt werden.
Die vierte Episode: PKM2 als extrazellulärer Signalkommunikator
Das Vorhandensein von extrazellulärem PKM2 eröffnet einen neuen Weg für die Untersuchung der biologischen Funktion von PKM2. Buschow et al. lieferte den ersten Beweis, dass PKM2 in B-Zell-Exosomen nachgewiesen werden konnte und als MHC-Klasse-II-assoziiertes Protein identifiziert wurde . Zwei nachfolgende Studien zeigten auch, dass PKM2 in Exosomen vorhanden ist, die von verschiedenen Zellen freigesetzt werden . Derzeit bieten mehrere öffentliche Datenbanken wie ExoCarta und EVpedia umfassende Informationen zu den Komponenten, einschließlich Proteinen, Lipiden und Nukleinsäuren extrazellulärer Vesikel in verschiedenen Spezies. Alle Daten bestätigen, dass PKM2 ein Paketprotein von Exosomen ist. Neuere Studien haben eindeutig eine kommunikative Rolle von Exosomen gezeigt, indem sie verschiedene Komponenten von Wirtszellen zu Empfängerzellen liefern . Es ist zu erwarten, dass PKM2 eine Rolle beim Zell-Zell-Übersprechen spielen kann.
Neue Erkenntnisse stützen diese Hypothese. Beispielsweise zeigte eine kürzlich durchgeführte Studie, dass zirkulierendes PKM2 im Blut das Tumorwachstum und die Angiogenese fördern kann, indem es das Wachstum, die Migration und die Matrixadhäsion von Endothelzellen erhöht . Eine andere Untersuchung zeigte auch, dass PKM2, das aus Darmkrebszellen ausgeschieden wird, über eine autokrine Stimulation wirken könnte, um die Zellmigration durch Aktivierung der PI3K / Akt- und Wnt / β-Catenin-Signalwege zu verbessern . Zusätzlich zu Krebszellen könnten Neutrophile an den Gewebeschädigungsstellen PKM2 freisetzen, um die Angiogenese und Wundheilung zu fördern . Unsere jüngste Studie zeigte auch, dass rekombinantes PKM2-Protein die Phosphorylierung und Aktivierung des epidermalen Wachstumsfaktorrezeptors (EGFR) induzieren kann . Darüber hinaus fanden wir heraus, dass die R339E-Mutante PKM2, die bevorzugt dimeres PKM2 bildete, EGFR signifikanter aktivierte als das tetramere PKM2. In: Keller et al. identifizierte 154 Proteine als potenzielle Substrate für PKM2 nach Behandlung von Hela-Zellen mit Succinyl-5-Aminoimidazol-4-carboxamid-1-Ribose-5′-phosphat (SAICAR), einem intrazellulären Metaboliten, der die Proteinkinaseaktivität von PKM2 stimulieren könnte . Sie fanden auch EGFR als PKM2-Substrat. Ihre Ergebnisse unterscheiden sich in zweierlei Hinsicht von unseren. Erstens werden die in unserer Studie aktivierten Signalwege durch extrazelluläres PKM2 ausgelöst, während die in ihrer Studie identifizierten molekularen Ziele potenzielle Substrate von intrazellulärem PKM2 sind. Zweitens Erhöhung der ERK1 / 2-Aktivität in unserer Studie wird durch EGFR-Aktivierung initiiert, während die ERK1 / 2-Aktivierung in ihrer Studie direkt durch den SAICAR / PKM2-Komplex stimuliert wird. Ein ähnliches Phänomen, das in beiden Studien beobachtet wurde, ist, dass die R339E-Mutante PKM2 Signalmoleküle signifikanter aktiviert als die Wildtyp-PKM2, was auf die unterschiedliche Rolle von dimerem und tetramerem PKM2 bei der Onkogenese hindeutet. Durch die Verwendung des Rezeptor-Tyrosinkinase-Arrays fanden wir heraus, dass extrazelluläres PKM2 nur begrenzte Wachstumsfaktorrezeptoren in Brustkrebszellen aktivierte (Daten nicht gezeigt). Derzeit ist die Selektivität der Rezeptoraktivierung durch extrazelluläres PKM2 unbekannt. Darüber hinaus ist auch nicht klar, warum die R339E-Mutante PKM2 bei der Aktivierung von EGFR wirksamer ist. Weitere Experimente sind erforderlich, um diese Fragen zu beantworten.
Eine weitere elegante Frage ist, ob freies PKM2 und vesikelverpackendes PKM2 eine ähnliche Wirkung bei der Förderung der Tumorentstehung zeigen (Abb. 2). Freies extrazelluläres PKM2 konnte die Plasmamembran nicht durchdringen und konnte nur intrazelluläre Signale über Zelloberflächenproteine wie Wachstumsfaktorrezeptoren aktivieren. Umgekehrt könnte vesikelverpackendes PKM2 von Krebs- und Stromazellen endozytiert werden, und das aus Vesikeln freigesetzte PKM2 könnte den Stoffwechsel und die Genexpression über intrazelluläre Mechanismen beeinflussen. The PKM2-null cells or mice will be useful to elucidate whether these two extracellular forms of PKM2 could act synergistically or antagonistically in tumorigenesis.

The potential tumor-promoting effect of extracellular PKM2
The fifth episode: unbeantwortete Diskrepanz von PKM2
Zusätzlich zu den oben diskutierten zellbasierten Daten wurde die onkogene Rolle von PKM2 auch nach der Erzeugung von PKM2-Knockout-Mäusen in Frage gestellt. In: Israelsen et al. generierte ein bedingtes Knockout-Mausmodell durch Löschen des PKM2-spezifischen Exons 10 . Überraschenderweise beschleunigte die Depletion von PKM2 die Tumorbildung, die durch den Verlust des Brca1-Gens bei Mäusen verursacht wurde, aber nicht abgeschwächt wurde. Diese Daten zeigten, dass PKM2 für die Proliferation von Krebszellen nicht erforderlich ist. Interessanterweise wurde die PKM1-Expression nur in nicht proliferierenden Tumorzellen nachgewiesen, was auf eine tumorsuppressive Rolle von PKM1 bei Brustkrebs hindeutet. Darüber hinaus haben PKM2-Knockout-Mäuse eine hohe Inzidenz, um nach einer langen Latenz aufgrund des Ungleichgewichts im Stoffwechsel spontan ein hepatozelluläres Karzinom zu entwickeln . Diese Ergebnisse widerlegen die Vorstellung, dass PKM2 in vivo eine onkogene Rolle spielt.
Die anhaltende Episode: Ist PKM2 ein Krebsbiomarker und ein Wirkstoffziel?
Obwohl die Ergebnisse des gentechnisch veränderten Mausmodells die tumorfördernde Aktivität von PKM2 nicht unterstützen, wird eine Überexpression von PKM2 allgemein bei menschlichen Krebserkrankungen gefunden und ist mit einem schlechten klinischen Ergebnis verbunden (Tabelle 1) . Zwei kürzlich durchgeführte Meta-Analyse-Studien unterstützten diese Schlussfolgerung ebenfalls. Wu et al. analysierte die Daten von 2812 Patienten mit soliden Tumoren des Verdauungssystems, die aus 16 Kohortenstudien gewonnen wurden, und stellte fest, dass eine Überexpression von PKM2 mit einem verringerten Gesamtüberleben bei Magenkrebs, Plattenepithelkarzinom der Speiseröhre, hepatozellulärem Karzinom, Gallenkrebs und Mundkrebs verbunden ist . PKM2 ist jedoch kein prognostischer Faktor für Bauchspeicheldrüsenkrebs. Dieser Befund stimmt nicht mit früheren Studien überein, die zeigen, dass der Anstieg von PKM2 ein Indikator für ein schlechtes Überleben von Bauchspeicheldrüsenkrebspatienten ist . Eine weitere Untersuchung umfasste 4796 Fälle aus 27 Einzelstudien, die zeigten, dass die PKM2-Hochregulation in gepoolten Daten mit einem schlechteren Gesamtüberleben, einem krankheitsfreien Überleben und einem rezidivfreien Überleben korreliert . Geschichtet nach Krebstyp sagt PKM2 jedoch kein schlechtes Überleben von Bauchspeicheldrüsenkrebs voraus. Insgesamt scheint PKM2 bei den meisten soliden Tumoren ein guter prognostischer Marker zu sein.
Im Gegenteil, die Verwendung von PKM2 als diagnostischer Faktor ist umstritten. Eine Proteomanalyse zeigte, dass PKM2 ein potenzieller diagnostischer Marker für den Nachweis von Lungenkrebs ist . Eine kürzlich durchgeführte Studie ergab jedoch, dass PKM2 aufgrund seiner geringen Spezifität kein guter diagnostischer Marker für Lungenkrebs ist . Ebenso ist es unwahrscheinlich, dass PKM2 allein ein nützlicher Marker für das Screening von Darmkrebs ist . Die Kombination mehrerer Marker könnte jedoch die Sensitivität und Spezifität für die Krebsdiagnose erhöhen .
Das therapeutische Potenzial von PKM2 ist ein faszinierendes Ereignis in der Krebsbehandlung. Von einer Seite wird erwartet, dass die Hemmung von PKM2 die Glykolyse hemmt, die Gentranskription beeinträchtigt und die Zellproliferation unterdrückt. Daher scheinen PKM2-Inhibitoren gute Kandidaten für die Entwicklung von Krebsmedikamenten zu sein. Durch die Verwendung von Bibliotheks-Screening, Vander Heiden et al. identifizierte drei neue Klassen von PKM2-Inhibitoren und zeigte, dass die wirksamste Verbindung die PKM2-Aktivität hemmte und den Tod von Krebszellen induzierte . Kürzlich haben Ning et al. gefunden, dass neue Naphthochinonderivate potente PKM2-Inhibitoren sind . Eine wirksame Verbindung 3 k unterdrückte die Proliferation mehrerer Krebszelllinien bei submikromolaren Konzentrationen, während sie auf normale Zellen nur eine geringe schädliche Wirkung zeigte. Von der anderen Seite kann die Aktivierung von PKM2 auch das Tumorwachstum hemmen. Da das PKM2-Dimer mit geringer Aktivität die Hauptisoform ist, die die Glykolyse im Zytoplasma und die Gentranskription im Zellkern in Krebszellen auslöst, können PKM2-Aktivatoren, die die Bildung von tetramerem PKM2 fördern können, die Glykolyse auf den Mitochondrienweg umschalten und den Kerneintritt reduzieren, um die Gentranskription abzuschwächen. Beide Effekte beeinträchtigen die metabolische Nachfrage und die wachstumsunterstützende Signalgebung, die zur Tumorregression führt. Zwei Pioneer-Studien identifizierten verschiedene PKM2-Aktivatoren und charakterisierten ihre Spezifität in vitro . Eine nachfolgende Studie zeigte, dass PKM2-Aktivatoren tatsächlich die Tetramerbildung förderten und das Tumorwachstum in vivo unterdrückten . Diese Ergebnisse deuteten darauf hin, dass PKM2-Aktivatoren vielversprechende Krebsmedikamente sein könnten.
Resistenz gegen Chemotherapie ist eine große Blockade für die Krebsbehandlung. Die Überwindung der inhärenten Chemoresistenz von Krebszellen ist für die klinische Forschung dringend erforderlich. Die Rolle von PKM2 bei der Chemoresistenz von Krebszellen wurde aufgedeckt, und es wurde gezeigt, dass das Targeting von PKM2 chemoresistente Krebszellen re-sensibilisiert. Eine kürzlich durchgeführte Studie zeigte, dass CD44 mit PKM2 interagiert und die PKM2-Aktivität durch zunehmende Tyr105-Phosphorylierung von PKM2 unterdrückt. Die CD44-Ablation induzierte den Wechsel von der aeroben Glykolyse zur mitochondrialen Atmung und erhöhte die Produktion reaktiver Sauerstoffspezies (ROS), was zur Verbesserung der Cisplatinempfindlichkeit in Darmkrebszellen führte . Es wurde gezeigt, dass die Hemmung der PKM2-Aktivität die Glykolyse unterdrückt und die Cisplatinresistenz überwindet. Daher kann die Kombination von Cisplatin und PKM2-Inhibitoren eine wirksame Strategie für die Chemotherapie sein (Abb. 3). Es wurde auch berichtet, dass PKM2 an der Regulation der Gemcitabin-Resistenz in Bauchspeicheldrüsenkrebszellen beteiligt ist. In: Kim et al. zeigte, dass PKM2 die Gemcitabin-Resistenz durch negativ regulierende p38-vermittelte p53-Phosphorylierung fördert, wodurch die Transkriptionsaktivität von p53 verringert und die Expression proapoptotischer Gene unterdrückt wird . Darüber hinaus erhöht die ektopische Expression von R399E-PKM2, das vorzugsweise dimeres PKM2 bildet, die Resistenz von Bauchspeicheldrüsenkrebszellen gegenüber Gemcitabin. Diese Evidenz legt nahe, dass PKM2 über einen nicht-metabolischen Mechanismus zur Induktion von Arzneimittelresistenzen beiträgt. Da dimeres PKM2 hauptsächlich als Transkriptionsregulator in Krebszellen wirkt, können PKM2-Aktivatoren, die die Bildung von tetramerem PKM2 fördern, bei der Unterdrückung der nichtmetabolischen Funktion von PKM2 und der Verringerung der Resistenz gegen Gemcitabin in Pankreaskrebszellen wirksam sein. Es wurde auch gezeigt, dass nukleares PKM2 zur Resistenz von beiträgt EGFR-Inhibitor bei Darmkrebs und Lungenkrebs . In: Li et al. zeigte, dass die nukleare PKM2-vermittelte STAT3-Phosphorylierung die Empfindlichkeit von Darmkrebszellen gegenüber Gefitinib verringert und die Störung der Wechselwirkung von nuklearem PKM2 und STAT3 die Gefitinib-Empfindlichkeit in den Zellen wiederherstellt . In jüngerer Zeit zeigte eine andere Studie, dass PKM2 in den Zellkern transloziert und bei Stimulation des Wachstumsfaktors mit Poly-ADP-Ribose (PAR) interagiert. Die PAR-bindende Aktivität von PKM2 ist entscheidend für die Kernretention und Gentranskription von PKM2 und ist wichtig für die Förderung der Zellproliferation und des Tumorwachstums. Sie kamen zu dem Schluss, dass die Hemmung der PKM2-Kernfunktion die Resistenz von EGFR-mutierten Krebszellen überwinden kann . Beide Studien wiesen auf eine entscheidende Rolle von nuklearem PKM2 bei der Vermittlung von Arzneimittelresistenzen bei Krebs hin und schlugen vor, dass das Targeting von nuklearem PKM2 eine vielversprechende Strategie zur Überwindung der Resistenz sein könnte.

PKM2 trägt zur Proliferation und Arzneimittelresistenz in Krebszellen bei