La piruvato quinasa M2 alimenta múltiples aspectos de las células cancerosas: desde el metabolismo celular, la regulación transcripcional hasta la señalización extracelular
El análisis bioquímico al caracterizar la actividad enzimática que cataliza la formación de lactato a partir de glucosa en lisados celulares reveló la primera vía metabólica intracelular, la vía glicolítica. A partir de la purificación de fracciones que contenían actividad glucolítica, varios investigadores pioneros contribuyeron a la identificación de enzimas que participan en cada paso de la vía . Estos resultados construyen nuestro concepto moderno en el intercambio de respiración aeróbica y anaeróbica y producción de energía bajo diversas circunstancias fisiológicas y patológicas.
La existencia de una enzima que catalizaba la producción de ATP mediante la transferencia de un grupo fosfato de PEP a ADP en el hígado se informó por primera vez en 1934 . El aislamiento posterior de la enzima, conocida posteriormente como piruvato quinasa (PK), demostró diferencias en la distribución tisular y en la cinética catalítica, lo que sugiere que esta enzima puede tener diferentes isoformas . Entre 1982 y 1984, se clonaron varios genes PK a partir de levadura, pollo y rata . El estudio funcional de la PKM2 se inició mediante la identificación de un gen candidato en ratones a principios de la década de 1980 . Más tarde, Noguchi et al. demostró que dos isoformas de PK (PKM1 y PKM2) están codificadas por el mismo gen PKM a través de empalmes alternativos . En humanos, las isoformas PKM también se producen a través de un mecanismo de empalme similar al incluir los exones 9 y 10 en el ARNm PKM1 y PKM2 por separado .
Varios hallazgos llamaron la atención de los investigadores sobre el papel potencial de la PKM2 en la tumorogénesis. En primer lugar, PKM2 es la isoforma embrionaria que se expresa altamente durante el desarrollo animal. Su transcripción se atenúa en varios tejidos adultos mientras se reactiva en tumores . En segundo lugar, el estudio de la abundancia relativa de PKM1 y PKM2 en tejidos normales y tumorales demostró un cambio de la isoforma PKM1 a la isoforma PKM2 en varios cánceres, como el carcinoma hepatocelular . En tercer lugar, el cambio de empalme de ARNm de PKM1 a PKM2 se ve reforzado por el oncogén c-Myc, lo que sugiere que las células cancerosas participan activamente en este cambio para adaptarse a sus necesidades de proliferación y metabolismo . En cuarto lugar, la modulación de la actividad PKM2 por activadores o inhibidores afecta el crecimiento tumoral in vivo .
- El primer episodio: PKM2 como enzima metabólica en el citoplasma
- El segundo episodio: PKM2 como modulador de señalización en el citoplasma
- El tercer episodio: PKM2 como regulador transcripcional en el núcleo
- El cuarto episodio: PKM2 como comunicador de señalización extracelular
- The fifth episode: discrepancia sin respuesta de PKM2
- El episodio continuo: ¿es el PKM2 un biomarcador del cáncer y un objetivo farmacológico?
El primer episodio: PKM2 como enzima metabólica en el citoplasma
Dado que el papel de la PKM2 en el control metabólico de la glucólisis en células cancerosas se ha revisado ampliamente , aquí solo resumimos tres diferencias cruciales entre la catálisis mediada por PKM1 y PKM2 y el metabolismo celular. La primera diferencia es la interacción de subunidades. Tanto PKM1 como PKM2 son proteínas tetraméricas formadas por cuatro subunidades idénticas. Cada subunidad (o monómero) contiene cuatro dominios estructurales, incluidos A, B, C y N-terminal. El monómero primero se dimeriza y luego dos dímeros interactúan a través de la interfaz dímero-dímero orquestada por el dominio C del monómero para formar un tetrámero. Debido a que PKM1 y PKM2 incluyen diferentes exones en sus ARNm, esto cambia los aminoácidos codificados en el dominio C y altera la estabilidad del tetrámero. Bajo condición fisiológica, PKM1 se organiza constitutivamente como un tetrámero, mientras que PKM2 puede existir en tetrámero o dímero. La segunda diferencia es la regulación alostérica. Dependiendo de las concentraciones intracelulares de pequeñas moléculas y metabolitos, la actividad de PKM1 y PKM2 puede regularse de forma diferencial. Uno de los reguladores alostéricos más conocidos es la fructosa-1,6-bisfosfato (FBP). Este intermediario glucolítico se une directamente a la PKM2 y aumenta la afinidad de la PKM2 por la PEP . Por el contrario, la PPF no afecta significativamente la actividad de PKM1. Además de la FBP, se ha informado que otros metabolitos, aminoácidos y moléculas pequeñas afectan la actividad de la PKM2 (Fig. 1). Sin embargo, la concentración requerida para la activación o inhibición es alta y el efecto modulador es modesto. Todavía no está claro si estas pequeñas moléculas desempeñan un papel importante en el control de la actividad PKM en circunstancias fisiológicas. La actividad de la PKM2 también está regulada por la modificación post-traslacional, como la fosforilación, acetilación y oxidación, que favorecen la baja actividad de la PKM2 dimérica (Fig. 1). La tercera diferencia es la producción de energía y la utilización intermedia. Dado que PKM1 existe constitutivamente como el tetrámero activo, la función biológica principal de esta isoforma es la generación de ATP para suministrar energía celular. Sin embargo, el PKM2, además de producir ATP, puede cambiar a la forma dimérica menos activa para generar varios intermedios glicolíticos que se pueden usar como bloques de construcción para la biosíntesis de aminoácidos, lípidos y nucleótidos.
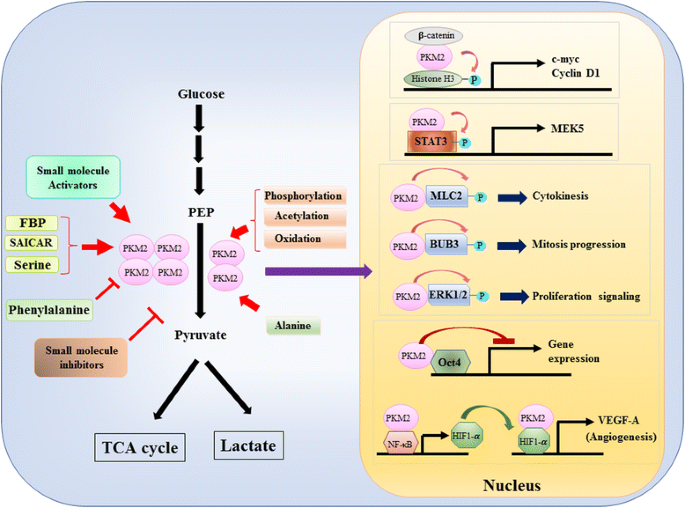
Modulación de la actividad de PKM2 por moléculas fisiológicas y modificación postraduccional y el papel potencial de PKM2 en la regulación transcripcional
El segundo episodio: PKM2 como modulador de señalización en el citoplasma
Además de funcionar como enzima glucolítica, se propone que la PKM2 participe en procesos más celulares debido a la identificación de proteínas que interactúan en el citoplasma. Por ejemplo, se demostró que la PKM2 es una proteína que interactúa con varias tirosina quinasas, incluida la A-Raf, la cinasa de fusión de la región del grupo de punto de ruptura-Abelson (BCR-ABL), el receptor del factor de crecimiento de fibroblastos 1 (FGFR1), etc. . Se ha demostrado que estas parejas de unión modulan el cambio dimérico/tetramérico de la PKM2 para alterar el metabolismo celular. Sin embargo, es posible que PKM2 pueda afectar recíprocamente la cinética catalítica, la unión al sustrato y la ubicación citoplasmática de estos socios de unión para modular la transducción de señales. El hallazgo de que PKM2 es una proteína de unión a fósforo-tirosina refuerza esta posibilidad porque muchos mediadores de señalización intracelular pueden unirse a residuos de fósforo-tirosina para ensamblar complejos proteicos específicos para la transmisión de señales . Hasta la fecha, la lista de socios de enlace PKM2 crece continuamente. Destacamos a varios miembros nuevos y discutimos sus implicaciones biológicas aquí. Mukheriee et al. se demostró que la PKM2 podría unirse con el HuR, una proteína de unión al ARN que desempeña un papel importante en el control de la estabilidad del ARNm y la eficiencia de la traducción, para promover la progresión del ciclo celular y la proliferación de células de glioma . Curiosamente, otra proteína de unión a ARN, tristetraprolina, que podría unirse a varios ARNm a través del elemento rico en AU en la región 3′-no traducible (3′-UTR), también se encontró que era un socio que interactuaba con PKM2, y PKM2 indujo fosforilación y degradación de tristetraprolina para modular el crecimiento del cáncer de mama . Estos dos estudios implican una posible función de control traslacional de PKM2. Recientemente, Liang et al. se identificó a la proteína antiapoptótica Bcl2 como un nuevo socio de PKM2 . Demostraron que el estrés oxidativo indujo la translocación de PKM2 a mitocondrias donde fosforiló y estabilizó Bcl2 al prevenir su degradación a través de la vía dependiente de la ubiquitinación. Estos datos sugirieron que la PKM2 ayuda a las células cancerosas a adaptarse al estrés oxidativo provocado por el cambio metabólico intracelular o el insulto extracelular.
El tercer episodio: PKM2 como regulador transcripcional en el núcleo
Un papel nuclear de PKM2 en la regulación de la transcripción génica o la modificación epigenética fue sugerido en primer lugar por el hallazgo de que PKM2 unido a β-catenina fosforilada Y333, y el complejo β-catenina-PKM2 fue reclutado a los nucleosomas para fosforilar la histona H3 a la treonina 11 (Fig. 1). Esta fosforilación aumentó posteriormente la acetilación de la histona H3 que condujo a la regulación ascendente de los genes diana de β-catenina. Otro factor de transcripción fosforilado directamente por PKM2 es el transductor de señal y activador de transcripción 3 (STAT3). La fosforilación de STAT3 mediada por PKM2 en la tirosina 705 aumentó la actividad de STAT3 para regular al alza la expresión de la proteína quinasa quinasa 5 activada por mitógenos (MEK5). Además de los factores de transcripción, se ha demostrado que PKM2 fosforila la cadena ligera de miosina 2 (MLC2), BUB3 y la quinasa regulada por señales extracelulares 1 y 2 (ERK1 y ERK2) . Curiosamente, el PKM2 también actúa de manera independiente de la fosforilación para afectar la expresión génica. Por ejemplo, se ha encontrado que la PKM2 se une a Oct4, uno de los factores de transcripción maestros que controlan la autorrenovación de las células madre e inhiben la transcripción mediada por Oct4 . PKM2 también puede mejorar la angiogénesis tumoral al interactuar con NF-kB y HIF-1α en el núcleo y activar la expresión del gen diana de HIF-1α VEGF-A. En consecuencia, el aumento de la secreción de VEGF-A aumenta la formación de vasos sanguíneos, lo que contribuye al crecimiento tumoral . Aunque estos estudios sugirieron fuertemente la localización nuclear y la función de la proteína quinasa de la PKM2 en diversas circunstancias fisiológicas y patológicas, sin embargo, la importancia de la expresión génica mediada por la PKM2 nuclear ha sido cuestionada por estudios que utilizan células eliminatorias de la PKM2. Mediante el uso de fibroblastos embrionarios de ratón nulos con PEP y PKM2 marcados, Hosios et al. demostró que la fosforilación dependiente de la PEP no es un evento común en las células y que la reacción no es catalizada por la PKM2 . La discrepancia de estos estudios está actualmente sin resolver y la actividad de la proteína quinasa de PKM2 necesita confirmación adicional.
El cuarto episodio: PKM2 como comunicador de señalización extracelular
La presencia de PKM2 extracelular abre una nueva vía para el estudio de la función biológica de la PKM2. Buschow et al. proporcionó la primera evidencia de que PKM2 se podía detectar en exosomas de células B y se identificó como una proteína asociada a MHC de clase II . Dos estudios posteriores también indicaron que existe PKM2 en exosomas liberados por varias células . Actualmente, varias bases de datos públicas como ExoCarta y EVpedia proporcionan información completa sobre los componentes, incluyendo proteínas, lípidos, ácidos nucleicos de vesículas extracelulares en diferentes especies. Todos los datos confirman que PKM2 es una proteína de paquete de exosomas. Estudios recientes han demostrado claramente el papel comunicativo de los exosomas al entregar diferentes componentes de las células huésped a las células receptoras . Es previsible que el PKM2 pueda desempeñar un papel en la diafonía celular-celular.
La evidencia emergente de hecho apoya esta hipótesis. Por ejemplo, un estudio reciente demostró que la PKM2 circulante en sangre puede promover el crecimiento tumoral y la angiogénesis al aumentar el crecimiento, la migración y la adhesión a la matriz de las células endoteliales . Otra investigación también mostró que la PKM2 secretada por células de cáncer de colon podría actuar a través de una estimulación autocrina para mejorar la migración celular mediante la activación de las vías PI3K/Akt y Wnt/β-catenina . Además de las células cancerosas, los neutrófilos en los sitios dañados de los tejidos podrían liberar PKM2 para promover la angiogénesis y la cicatrización de heridas . Nuestro estudio reciente también demostró que la proteína PKM2 recombinante podría inducir fosforilación y activación del receptor del factor de crecimiento epidérmico (EGFR) . Además, encontramos que el mutante PKM2 de R339E que formaba preferentemente PKM2 dimérico activaba el EGFR de manera más significativa que el PKM2 tetramérico. Keller et al. se identificaron 154 proteínas como sustratos potenciales para la PKM2 después del tratamiento de células Hela con succinil-5-aminoimidazol-4-carboxamida-1-ribosa-5′-fosfato (SAICAR), un metabolito intracelular que podría estimular la actividad de la proteína quinasa de la PKM2 . También encontraron EGFR como sustrato de PKM2. Sus resultados son diferentes de los nuestros en dos sentidos. En primer lugar, las vías de señalización activadas en nuestro estudio son provocadas por la PKM2 extracelular, mientras que las dianas moleculares identificadas en su estudio son sustratos potenciales de la PKM2 intracelular. En segundo lugar, el aumento de la actividad de ERK1/2 en nuestro estudio se inicia mediante la activación de EGFR, mientras que la activación de ERK1/2 en su estudio es estimulada directamente por el complejo SAICAR/PKM2. Un fenómeno similar observado en ambos estudios es que la PKM2 mutante R339E activa moléculas de señalización de manera más significativa que la PKM2 de tipo salvaje, lo que sugiere el papel distinto de la PKM2 dimérica y tetramérica en la oncogénesis. Mediante el uso de una matriz de receptores de tirosina quinasa, descubrimos que la PKM2 extracelular solo activaba receptores de factor de crecimiento limitado en células de cáncer de mama (no se muestran datos). Actualmente, la selectividad de la activación del receptor por PKM2 extracelular sigue siendo desconocida. Además, tampoco está claro por qué el mutante R339E PKM2 es más potente en la activación del EGFR. Se necesitan más experimentos para responder a estas preguntas.
Otra pregunta elegante que debe abordarse es si el PKM2 libre y el PKM2 envasado en vesículas exhiben un efecto similar en la promoción de la tumorogénesis (Fig. 2). El PKM2 extracelular libre no podía penetrar la membrana plasmática y solo podía activar la señalización intracelular a través de proteínas de la superficie celular como los receptores del factor de crecimiento. Por el contrario, la PKM2 empaquetadora de vesículas podría ser endocitosada por células cancerosas y estromas, y la PKM2 liberada de las vesículas podría afectar el metabolismo y la expresión génica a través del mecanismo intracelular. The PKM2-null cells or mice will be useful to elucidate whether these two extracellular forms of PKM2 could act synergistically or antagonistically in tumorigenesis.

The potential tumor-promoting effect of extracellular PKM2
The fifth episode: discrepancia sin respuesta de PKM2
Además de los datos basados en células discutidos anteriormente, el papel oncogénico de PKM2 también se ha cuestionado después de la generación de ratones knockout de PKM2. Israelsen et al. se generó un modelo de ratón knockout condicional eliminando el exón 10 específico de PKM2 . Sorprendentemente, el agotamiento de la PKM2 aceleró pero no atenuó la formación de tumores debido a la pérdida del gen Brca1 en ratones. Estos datos indicaron que la PKM2 no es necesaria para la proliferación de células cancerosas. Curiosamente, la expresión de PKM1 solo se detectó en células tumorales no proliferantes, lo que indica una función supresora tumoral de la PKM1 en el cáncer de mama. Además, los ratones knockout PKM2 tienen una alta incidencia de desarrollar carcinoma hepatocelular espontáneamente después de una larga latencia debido al desequilibrio en el metabolismo . Estos resultados contrastan con la noción de que el PKM2 desempeña un papel oncogénico in vivo.
El episodio continuo: ¿es el PKM2 un biomarcador del cáncer y un objetivo farmacológico?
Aunque los resultados del modelo de ratón de ingeniería genética no respaldan la actividad promotora tumoral de la PKM2, la sobreexpresión de la PKM2 se encuentra universalmente en los cánceres humanos y se relaciona con un desenlace clínico precario (Tabla 1) . Dos estudios recientes de metanálisis también respaldaron esta conclusión. Wu et al. se analizaron los datos de 2.812 pacientes con tumores sólidos del sistema digestivo obtenidos de 16 estudios de cohortes y se encontró que la sobreexpresión de PKM2 se relaciona con una reducción de la supervivencia general en cáncer gástrico, carcinoma de células escamosas de esófago, carcinoma hepatocelular, cáncer biliar y cáncer oral . Sin embargo, la PKM2 no es un factor pronóstico para el cáncer de páncreas. Este hallazgo no concuerda con estudios previos que mostraron que el aumento de PKM2 es un indicador de supervivencia precaria de los pacientes con cáncer de páncreas . Otra investigación incluyó 4.796 casos de 27 estudios individuales que demostraron que la regulación ascendente de la PKM2 se correlaciona con una supervivencia general peor, una supervivencia sin enfermedad y una supervivencia sin recidiva en los datos agrupados . Sin embargo, estratificada por tipo de cáncer, la PKM2 no predice una supervivencia precaria del cáncer de páncreas. En conjunto, la PKM2 parece ser un marcador pronóstico responsable en la mayoría de los tumores sólidos.
Por el contrario, el uso de la PKM2 como factor diagnóstico es controvertido. Un análisis proteómico demostró que la PKM2 es un marcador diagnóstico potencial para la detección de cáncer de pulmón . Sin embargo, un estudio reciente sugirió que la PKM2 no es un buen marcador diagnóstico para el cáncer de pulmón debido a la baja especificidad . Del mismo modo, es poco probable que la PKM2 por sí sola sea un marcador útil para la detección del cáncer de colon . Sin embargo, la combinación de múltiples marcadores podría aumentar la sensibilidad y la especificidad para el diagnóstico de cáncer .
El potencial terapéutico del PKM2 es un evento intrigante en el tratamiento del cáncer. Por un lado, se espera que la inhibición de PKM2 inhiba la glucólisis, perjudique la transcripción génica y suprima la proliferación celular. Por lo tanto, los inhibidores de PKM2 parecen ser buenos candidatos para el desarrollo de medicamentos anticancerosos. Mediante el uso de la detección de bibliotecas, Vander Heiden et al. se identificaron tres nuevas clases de inhibidores de la PKM2 y se observó que el compuesto más eficaz inhibía la actividad de la PKM2 e inducía la muerte de células cancerosas . Recientemente, Ning et al. se encontró que los nuevos derivados de la naftoquinona son potentes inhibidores de la PKM2 . Un compuesto eficaz 3 k suprimió la proliferación de múltiples líneas celulares cancerosas a concentraciones sub-micromolares, mientras que mostró poco efecto perjudicial sobre las células normales. Desde el otro lado, la activación de la PKM2 también puede inhibir el crecimiento tumoral. Debido a que el dímero PKM2 de baja actividad es la isoforma principal que desencadena la glucólisis en el citoplasma y la transcripción génica en el núcleo de las células cancerosas, los activadores de PKM2 que pueden promover la formación de PKM2 tetramérico pueden cambiar la glucólisis a la vía mitocondrial y reducir la entrada nuclear para atenuar la transcripción génica. Ambos efectos afectan la demanda metabólica y la señalización de apoyo al crecimiento que conduce a la regresión tumoral. Dos estudios pioneros identificaron varios activadores de PKM2 y caracterizaron su especificidad in vitro . Un estudio posterior demostró que los activadores de PKM2 de hecho promovieron la formación de tetrámeros y suprimieron el crecimiento tumoral in vivo . Estos resultados sugirieron que los activadores de PKM2 podrían ser medicamentos anticancerosos prometedores.
La resistencia a la quimioterapia es un bloqueo importante para el tratamiento del cáncer. La superación de la quimiorresistencia inherente de las células cancerosas es urgente para la investigación clínica. Se han revelado las funciones de la PKM2 en la resistencia a la quimiorresistencia de las células cancerosas, y se ha demostrado que la focalización de la PKM2 vuelve a sensibilizar a las células cancerosas quimiorresistentes. Un estudio reciente mostró que CD44 interactúa con PKM2 y suprime la actividad de PKM2 a través del aumento de la fosforilación Tyr105 de PKM2. La ablación de CD44 indujo el cambio de la glucólisis aeróbica a la respiración mitocondrial y el aumento de la producción de especies reactivas de oxígeno (ROS), lo que resultó en una mejora de la sensibilidad al cisplatino en las células cancerosas colorrectales . Se demostró que la inhibición de la actividad PKM2 suprime la glucólisis y supera la resistencia al cisplatino. Por lo tanto, la combinación de cisplatino e inhibidores de la PKM2 puede ser una estrategia eficaz para la quimioterapia (Fig. 3). También se ha notificado que la PKM2 participa en la regulación de la resistencia a la gemcitabina en las células cancerosas de páncreas. Kim et al. demostró que la PKM2 promueve la resistencia a la gemcitabina a través de la regulación negativa de la fosforilación de p53 mediada por p38, lo que reduce la actividad transcripcional de p53 y suprime la expresión de genes proapoptóticos . Además, la expresión ectópica de R399E-PKM2, que forma preferentemente PKM2 dimérico, aumenta la resistencia de las células cancerosas pancreáticas a la gemcitabina. Estas evidencias sugieren que la PKM2 contribuye a la inducción de resistencia a los medicamentos a través de un mecanismo no metabólico. Debido a que la PKM2 dimérica actúa principalmente como un regulador de la transcripción en las células cancerosas, los activadores de la PKM2 que promueven la formación de PKM2 tetramérico pueden ser eficaces para suprimir la función no metabólica de la PKM2 y reducir la resistencia a la gemcitabina en las células cancerosas de páncreas. También se ha demostrado que la PKM2 nuclear contribuye a la resistencia del inhibidor del EGFR en el cáncer colorrectal y el cáncer de pulmón . Li et al. mostró que la fosforilación de STAT3 mediada por PKM2 nuclear reduce la sensibilidad de las células cancerosas colorrectales al gefitinib y la interrupción de la interacción de PKM2 nuclear y STAT3 restauró la sensibilidad de gefitinib en las células . Más recientemente, otro estudio demostró que PKM2 se trasloca al núcleo e interactúa con poli-ADP ribosa (PAR) al estimular el factor de crecimiento. La actividad de unión a PAR de la PKM2 es crítica para la retención nuclear y la transcripción génica de la PKM2 y es importante para la promoción de la proliferación celular y el crecimiento tumoral. Concluyeron que la inhibición de la función nuclear PKM2 puede superar la resistencia de las células cancerosas mutadas en EGFR . Ambos estudios señalaron un papel crucial de la PKM2 nuclear en la mediación de la resistencia a los medicamentos en los cánceres y sugirieron que atacar la PKM2 nuclear podría ser una estrategia prometedora para anular la resistencia.

PKM2 contribuye a la proliferación y resistencia a los fármacos en las células de cáncer