La pyruvate kinase M2 alimente de multiples aspects des cellules cancéreuses: du métabolisme cellulaire, de la régulation transcriptionnelle à la signalisation extracellulaire
L’analyse biochimique en caractérisant l’activité enzymatique qui catalyse la formation de lactate à partir du glucose dans les lysats cellulaires a révélé la première voie métabolique intracellulaire, la voie glycolytique. À partir de la purification des fractions contenant une activité glycolytique, un certain nombre de chercheurs pionniers ont contribué à l’identification d’enzymes impliquées dans chaque étape de la voie. Ces résultats construisent notre concept moderne d’échange de respiration aérobie et anaérobie et de production d’énergie dans diverses circonstances physiologiques et pathologiques.
L’existence d’une enzyme qui catalysait la production d’ATP en transférant un groupe phosphate du PEP à l’ADP dans le foie a été signalée pour la première fois en 1934. L’isolement ultérieur de l’enzyme, connue plus tard sous le nom de pyruvate kinase (PK), a démontré des différences dans la distribution tissulaire et la cinétique catalytique suggérant que cette enzyme pourrait avoir des isoformes différentes. De 1982 à 1984, divers gènes PK ont été clonés à partir de levure, de poulet et de rat. L’étude fonctionnelle de PKM2 a été initiée par l’identification d’un gène candidat chez la souris au début des années 1980. Plus tard, Noguchi et coll. a montré que deux isoformes de PK (PKM1 et PKM2) sont codées par le même gène PKM via un épissage alternatif. Chez l’homme, les isoformes PKM sont également produites via un mécanisme d’épissage similaire en incluant les exons 9 et 10 dans l’ARNm PKM1 et PKM2 séparément.
Plusieurs résultats ont attiré l’attention des chercheurs sur le rôle potentiel de la PKM2 dans la tumorigénèse. Premièrement, PKM2 est l’isoforme embryonnaire qui s’exprime fortement au cours du développement animal. Sa transcription est atténuée dans un certain nombre de tissus adultes tandis qu’elle est réactivée dans les tumeurs. Deuxièmement, l’étude de l’abondance relative de PKM1 et de PKM2 dans les tissus normaux et tumoraux a démontré un passage de l’isoforme PKM1 à l’isoforme PKM2 dans divers cancers comme le carcinome hépatocellulaire. Troisièmement, le changement de l’épissage de l’ARNm de PKM1 à PKM2 est renforcé par l’oncogène c-Myc suggérant que les cellules cancéreuses participent activement à ce changement pour répondre à leurs besoins en prolifération et en métabolisme. Quatrièmement, la modulation de l’activité PKM2 par des activateurs ou des inhibiteurs affecte la croissance tumorale in vivo.
- Le premier épisode: PKM2 en tant qu’enzyme métabolique dans le cytoplasme
- Le deuxième épisode: PKM2 comme modulateur de signalisation dans le cytoplasme
- Le troisième épisode: PKM2 en tant que régulateur transcriptionnel dans le noyau
- Le quatrième épisode : PKM2 en tant que communicateur de signalisation extracellulaire
- The fifth episode: divergence sans réponse de PKM2
- L’épisode qui se poursuit : la PKM2 est-elle un biomarqueur du cancer et une cible médicamenteuse?
Le premier épisode: PKM2 en tant qu’enzyme métabolique dans le cytoplasme
Étant donné que le rôle de PKM2 dans le contrôle métabolique de la glycolyse dans les cellules cancéreuses a été largement examiné, nous ne résumons ici que trois différences cruciales entre la catalyse médiée par PKM1 et PKM2 et le métabolisme cellulaire. La première différence est l’interaction des sous-unités. PKM1 et PKM2 sont toutes deux des protéines tétramériques formées de quatre sous-unités identiques. Chaque sous-unité (ou monomère) contient quatre domaines structuraux, y compris le domaine A, B, C et N-terminal. Le monomère se dimérise d’abord ensemble, puis deux dimères interagissent via l’interface dimère-dimère orchestrée par le domaine C du monomère pour former un tétramère. Parce que PKM1 et PKM2 incluent différents exons dans leurs ARNM, cela modifie les acides aminés codés dans le domaine C et modifie la stabilité du tétramère. Dans des conditions physiologiques, PKM1 s’organise constitutivement comme un tétramère tandis que PKM2 peut exister dans un tétramère ou un dimère. La deuxième différence est la régulation allostérique. En fonction des concentrations intracellulaires de petites molécules et de métabolites, l’activité de PKM1 et de PKM2 peut être régulée de manière différentielle. L’un des régulateurs allostériques les plus connus est le fructose-1,6-bisphosphate (FBP). Cet intermédiaire glycolytique lie directement PKM2 et augmente l’affinité de PKM2 pour la PEP. Au contraire, FBP n’affecte pas de manière significative l’activité PKM1. En plus de la FBP, d’autres métabolites, acides aminés et petites molécules ont été rapportés pour affecter l’activité de la PKM2 (Fig. 1). Cependant, la concentration requise pour l’activation ou l’inhibition est élevée et l’effet modulateur est modeste. On ne sait toujours pas si ces petites molécules jouent un rôle important dans le contrôle de l’activité PKM dans des circonstances physiologiques. L’activité de la PKM2 est également régulée par des modifications post-traductionnelles, telles que la phosphorylation, l’acétylation et l’oxydation, qui favorisent la faible activité de la PKM2 dimérique (Fig. 1). La troisième différence concerne la production d’énergie et l’utilisation intermédiaire. Puisque PKM1 existe constitutivement comme tétramère actif, la fonction biologique principale de cette isoforme est la génération d’ATP pour fournir de l’énergie cellulaire. Cependant, PKM2, en plus de produire de l’ATP, peut passer à la forme dimérique moins active pour générer plusieurs intermédiaires glycolytiques qui peuvent être utilisés comme blocs de construction pour la biosynthèse des acides aminés, des lipides et des nucléotides.
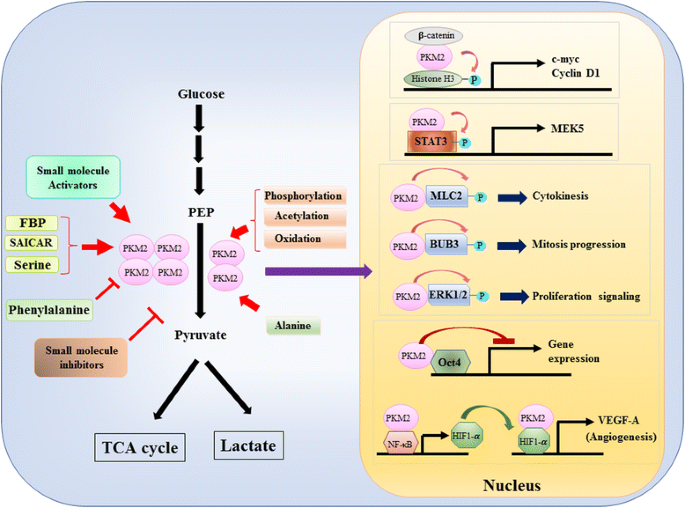
Modulation de l’activité de PKM2 par des molécules physiologiques et modification post-traductionnelle et rôle potentiel de PKM2 dans la régulation transcriptionnelle
Le deuxième épisode: PKM2 comme modulateur de signalisation dans le cytoplasme
En plus de fonctionner comme enzyme glycolytique, il est proposé que PKM2 s’implique dans des processus plus cellulaires en raison de l’identification de protéines en interaction dans le cytoplasme. Par exemple, PKM2 s’est avéré être une protéine interagissant avec plusieurs tyrosines kinases, y compris l’A-Raf, la kinase de fusion de la région de cluster de point de rupture – Abelson (BCR-ABL), le récepteur du facteur de croissance des fibroblastes 1 (FGFR1), etc. . Il a été démontré que ces partenaires de liaison modulent le changement dimérique / tétramérique de PKM2 pour modifier le métabolisme cellulaire. Cependant, il est possible que PKM2 affecte réciproquement la cinétique catalytique, la liaison au substrat et la localisation cytoplasmique de ces partenaires de liaison pour moduler la transduction du signal. La découverte que PKM2 est une protéine de liaison phosphor-tyrosine renforce cette possibilité car de nombreux médiateurs de signalisation intracellulaires peuvent se lier au résidu phosphor-tyrosine pour assembler des complexes protéiques spécifiques pour la transmission du signal. À ce jour, la liste des partenaires de liaison PKM2 ne cesse de s’allonger. Nous mettons en évidence plusieurs nouveaux membres et discutons de leur implication biologique ici. Mukheriee et al. a démontré que PKM2 pouvait se lier à HuR, une protéine de liaison à l’ARN qui joue un rôle important dans le contrôle de la stabilité de l’ARNm et de l’efficacité de la traduction, pour favoriser la progression du cycle cellulaire et la prolifération des cellules du gliome. Il est intéressant de noter qu’une autre protéine de liaison à l’ARN, la tristétrapoline, qui pourrait lier un certain nombre d’ARNm via l’élément riche en AU dans la région 3′-non traductionnelle (3′-UTR), s’est également avérée être un partenaire d’interaction PKM2, et PKM2 a induit la phosphorylation et la dégradation de la tristétrapoline pour moduler la croissance du cancer du sein. Ces deux études impliquent une fonction de contrôle translationnel potentielle de PKM2. Récemment, Liang et al. a identifié la protéine anti-apoptotique Bcl2 comme un nouveau partenaire PKM2. Ils ont démontré que le stress oxydatif induisait la translocation de la PKM2 dans les mitochondries où elle phosphorylait et stabilisait la Bcl2 en empêchant sa dégradation par voie dépendante de l’ubiquitination. Ces données suggèrent que la PKM2 aide les cellules cancéreuses à adapter le stress oxydatif provoqué par un changement métabolique intracellulaire ou une insulte extracellulaire.
Le troisième épisode: PKM2 en tant que régulateur transcriptionnel dans le noyau
Un rôle nucléaire de PKM2 dans la régulation de la transcription génique ou de la modification épigénétique a d’abord été suggéré par la découverte que PKM2 se liait à la β-caténine phosphorlatée par l’Y333, et que le complexe β-caténine-PKM2 était recruté dans les nucléosomes pour phosphoryler l’histone H3 à la thréonine 11 (Fig. 1). Cette phosphorylation a ensuite augmenté l’acétylation de l’histone H3, ce qui a conduit à une régulation à la hausse des gènes cibles de la β-caténine. Un autre facteur de transcription directement phosphorylé par PKM2 est le transducteur de signal et l’activateur de la transcription 3 (STAT3). La phosphorylation médiée par la PKM2 de la STAT3 à la tyrosine 705 a amélioré l’activité de la STAT3 pour réguler à la hausse l’expression de la protéine kinase kinase 5 activée par le mitogène (MEK5). Outre les facteurs de transcription, il a été démontré que PKM2 phosphorylait la chaîne légère de myosine 2 (MLC2), la BUB3 et la kinase 1 et 2 régulée par le signal extracellulaire (ERK1 et ERK2). Fait intéressant, PKM2 agit également de manière indépendante de la phosphorylation pour affecter l’expression des gènes. Par exemple, il a été constaté que PKM2 se lie à Oct4, l’un des facteurs de transcription principaux qui contrôlent l’auto-renouvellement des cellules souches et inhibent la transcription médiée par Oct4. PKM2 peut également améliorer l’angiogenèse tumorale en interagissant avec NF-kB et HIF-1α dans le noyau et en activant l’expression du gène cible HIF-1α VEGF-A. Par conséquent, une sécrétion accrue de VEGF-A stimule la formation de vaisseaux sanguins qui contribue à la croissance tumorale. Bien que ces études aient fortement suggéré la localisation nucléaire et la fonction de la protéine kinase de PKM2 dans diverses circonstances physiologiques et pathologiques, l’importance de l’expression génique médiée par PKM2 nucléaire a été remise en question par des études utilisant des cellules knockout PKM2. En utilisant des fibroblastes embryonnaires de souris marqués PEP et PKM2-null, Hosios et al. a montré que la phosphorylation PEP-dépendante n’est pas un événement courant dans les cellules et que la réaction n’est pas catalysée par la PKM2. La divergence de ces études n’est actuellement pas résolue et l’activité de la protéine kinase de PKM2 doit être confirmée.
Le quatrième épisode : PKM2 en tant que communicateur de signalisation extracellulaire
La présence de PKM2 extracellulaire ouvre une nouvelle voie pour l’étude de la fonction biologique de PKM2. Buschow et coll. a fourni la première preuve que la PKM2 pouvait être détectée dans les exosomes de cellules B et a été identifiée comme une protéine associée au CMH de classe II. Deux études ultérieures ont également indiqué que la PKM2 existe dans les exosomes libérés par diverses cellules. Actuellement, plusieurs bases de données publiques comme ExoCarta et EVpedia fournissent des informations complètes sur les composants, y compris les protéines, les lipides, les acides nucléiques des vésicules extracellulaires de différentes espèces. Toutes les données confirment que PKM2 est une protéine de paquet d’exosomes. Des études récentes ont clairement démontré un rôle communicatif des exosomes en délivrant différents composants des cellules hôtes aux cellules receveuses. Il est probable que PKM2 puisse jouer un rôle dans la diaphonie cellule-cellule.
Des preuves émergentes soutiennent en effet cette hypothèse. Par exemple, une étude récente a démontré que la PKM2 circulante dans le sang peut favoriser la croissance tumorale et l’angiogenèse en augmentant la croissance, la migration et l’adhésion matricielle des cellules endothéliales. Une autre étude a également montré que la PKM2 sécrétée par les cellules cancéreuses du côlon pouvait agir via une stimulation autocrine pour améliorer la migration cellulaire en activant les voies PI3K / Akt et Wnt / β-caténine. En plus des cellules cancéreuses, les neutrophiles aux sites des lésions tissulaires pourraient libérer de la PKM2 pour favoriser l’angiogenèse et la cicatrisation des plaies. Notre étude récente a également démontré que la protéine PKM2 recombinante pouvait induire la phosphorylation et l’activation du récepteur du facteur de croissance épidermique (EGFR). De plus, nous avons trouvé que le PKM2 mutant R339E qui formait préférentiellement du PKM2 dimérique activait l’EGFR de manière plus significative que le PKM2 tétramérique. Keller et coll. 154 protéines ont été identifiées comme substrats potentiels de la PKM2 après traitement des cellules Hela avec du succinyl-5-aminoimidazole-4-carboxamide-1-ribose-5′-phosphate (SAICAR), un métabolite intracellulaire qui pourrait stimuler l’activité de la protéine kinase de la PKM2. Ils ont également trouvé l’EGFR comme substrat PKM2. Leurs résultats sont différents des nôtres de deux manières. Premièrement, les voies de signalisation activées dans notre étude sont déclenchées par la PKM2 extracellulaire tandis que les cibles moléculaires identifiées dans leur étude sont des substrats potentiels de la PKM2 intracellulaire. Deuxièmement, l’augmentation de l’activité ERK1 / 2 dans notre étude est initiée par l’activation de l’EGFR tandis que l’activation ERK1 / 2 dans leur étude est directement stimulée par le complexe SAICAR / PKM2. Un phénomène similaire observé dans les deux études est que le PKM2 mutant du R339E active les molécules de signalisation de manière plus significative que le PKM2 de type sauvage suggérant le rôle distinct du PKM2 dimérique et tétramérique dans l’oncogenèse. En utilisant un réseau de récepteurs tyrosine kinase, nous avons constaté que la PKM2 extracellulaire n’activait que des récepteurs à facteur de croissance limité dans les cellules cancéreuses du sein (données non montrées). Actuellement, la sélectivité de l’activation des récepteurs par la PKM2 extracellulaire reste inconnue. De plus, on ne sait pas non plus pourquoi le PKM2 mutant R339E est plus puissant dans l’activation de l’EGFR. D’autres expériences sont nécessaires pour répondre à ces questions.
Une autre question élégante à aborder est de savoir si la PKM2 libre et la PKM2 contenant des vésicules ont un effet similaire sur la promotion de la tumorigénèse (Fig. 2). La PKM2 extracellulaire libre ne pouvait pas pénétrer la membrane plasmique et ne pouvait activer la signalisation intracellulaire que par l’intermédiaire de protéines de surface cellulaire telles que les récepteurs de facteurs de croissance. Inversement, le PKM2 empaquetant les vésicules pourrait être endocyté par les cellules cancéreuses et stroma, et le PKM2 libéré par les vésicules pourrait affecter le métabolisme et l’expression des gènes via un mécanisme intracellulaire. The PKM2-null cells or mice will be useful to elucidate whether these two extracellular forms of PKM2 could act synergistically or antagonistically in tumorigenesis.

The potential tumor-promoting effect of extracellular PKM2
The fifth episode: divergence sans réponse de PKM2
En plus des données cellulaires discutées ci-dessus, le rôle oncogène de PKM2 a également été contesté après la génération de souris knockout PKM2. Israelsen et coll. génération d’un modèle de souris knockout conditionnel en supprimant l’exon 10 spécifique à PKM2. Étonnamment, la déplétion de PKM2 a accéléré mais non atténué la formation de tumeurs entraînée par la perte du gène Brca1 chez la souris. Ces données indiquent que la PKM2 n’est pas nécessaire à la prolifération des cellules cancéreuses. Fait intéressant, l’expression de PKM1 n’a été détectée que dans des cellules tumorales non proliférantes suggérant un rôle suppresseur de tumeur de PKM1 dans le cancer du sein. De plus, les souris knockout PKM2 ont une incidence élevée de développer spontanément un carcinome hépatocellulaire après une longue latence due au déséquilibre du métabolisme. Ces résultats vont à l’encontre de l’idée que la PKM2 joue un rôle oncogène in vivo.
L’épisode qui se poursuit : la PKM2 est-elle un biomarqueur du cancer et une cible médicamenteuse?
Bien que les résultats du modèle murin d’ingénierie génétique ne soutiennent pas l’activité tumorale de PKM2, la surexpression de PKM2 est universellement présente dans les cancers humains et est associée à de mauvais résultats cliniques (tableau 1). Deux études récentes de méta-analyse ont également étayé cette conclusion. Wu et coll. a analysé les données de 2812 patients atteints de tumeurs solides du système digestif obtenues à partir de 16 études de cohorte et a constaté que la surexpression de PKM2 est associée à une survie globale réduite dans le cancer gastrique, le carcinome épidermoïde de l’œsophage, le carcinome hépatocellulaire, le cancer biliaire et le cancer buccal. Cependant, la PKM2 n’est pas un facteur pronostique pour le cancer du pancréas. Cette constatation n’est pas cohérente avec les études précédentes montrant que l’augmentation de la PKM2 est un indicateur de faible survie des patients atteints de cancer du pancréas. Une autre enquête a inclus 4796 cas de 27 études individuelles démontrant que la régulation à la hausse de la PKM2 est corrélée à une survie globale plus mauvaise, à une survie sans maladie et à une survie sans récidive dans les données regroupées. Cependant, stratifiée par type de cancer, la PKM2 ne prédit pas une faible survie du cancer du pancréas. Collectivement, PKM2 semble être un marqueur pronostique responsable dans la plupart des tumeurs solides.
Au contraire, l’utilisation de la PKM2 comme facteur diagnostique est controversée. Une analyse protéomique a démontré que la PKM2 est un marqueur diagnostique potentiel pour la détection du cancer du poumon. Cependant, une étude récente a suggéré que la PKM2 n’est pas un bon marqueur diagnostique du cancer du poumon en raison de sa faible spécificité. De même, il est peu probable que la PKM2 seule soit un marqueur utile pour le dépistage du cancer du côlon. Cependant, la combinaison de plusieurs marqueurs pourrait augmenter la sensibilité et la spécificité pour le diagnostic du cancer.
Le potentiel thérapeutique de la PKM2 est un événement intrigant dans le traitement du cancer. D’un côté, l’inhibition de la PKM2 devrait inhiber la glycolyse, altérer la transcription des gènes et supprimer la prolifération cellulaire. Par conséquent, les inhibiteurs de la PKM2 semblent être de bons candidats pour le développement de médicaments anticancéreux. En utilisant le filtrage de bibliothèque, Vander Heiden et al. a identifié trois nouvelles classes d’inhibiteurs de la PKM2 et a montré que le composé le plus efficace inhibait l’activité de la PKM2 et induisait la mort des cellules cancéreuses. Récemment, Ning et al. a constaté que les nouveaux dérivés de la naphtoquinone sont de puissants inhibiteurs de la PKM2. Un composé efficace 3 k a supprimé la prolifération de lignées cellulaires cancéreuses multiples à des concentrations sub-micromolaires alors qu’il a montré peu d’effet néfaste sur les cellules normales. De l’autre côté, l’activation de PKM2 peut également inhiber la croissance tumorale. Étant donné que le dimère PKM2 à faible activité est l’isoforme principale qui déclenche la glycolyse dans le cytoplasme et la transcription des gènes dans le noyau des cellules cancéreuses, les activateurs PKM2 qui peuvent favoriser la formation de PKM2 tétramérique peuvent basculer la glycolyse vers la voie mitochondriale et réduire l’entrée nucléaire pour atténuer la transcription des gènes. Les deux effets altèrent la demande métabolique et la signalisation de soutien à la croissance qui conduit à la régression tumorale. Deux études pionnières ont identifié divers activateurs de PKM2 et ont caractérisé leur spécificité in vitro. Une étude ultérieure a démontré que les activateurs de PKM2 favorisaient en effet la formation de tétramères et supprimaient la croissance tumorale in vivo. Ces résultats suggèrent que les activateurs de PKM2 pourraient être des médicaments anticancéreux prometteurs.
La résistance à la chimiothérapie est un blocage majeur pour le traitement du cancer. Surmonter la chimiorésistance inhérente aux cellules cancéreuses est urgent pour la recherche clinique. Les rôles de la PKM2 dans la chimiorésistance des cellules cancéreuses ont été révélés, et il a été démontré que le ciblage de la PKM2 re-sensibilise les cellules cancéreuses chimiorésistantes. Une étude récente a montré que CD44 interagit avec PKM2 et supprime l’activité PKM2 via une augmentation de la phosphorylation Tyr105 de PKM2. L’ablation de CD44 a induit le passage de la glycolyse aérobie à la respiration mitochondriale et l’augmentation de la production d’espèces réactives de l’oxygène (ROS), ce qui a entraîné l’amélioration de la sensibilité du cisplatine dans les cellules cancéreuses colorectales. L’inhibition de l’activité PKM2 a été démontrée pour supprimer la glycolyse et vaincre la résistance au cisplatine. Par conséquent, l’association du cisplatine et des inhibiteurs de la PKM2 peut être une stratégie efficace pour la chimiothérapie (Fig. 3). Il a également été rapporté que la PKM2 participe à la régulation de la résistance à la gemcitabine dans les cellules cancéreuses du pancréas. Kim et coll. a montré que la PKM2 favorise la résistance à la gemcitabine en régulant négativement la phosphorylation de la p53 médiée par la p38, réduisant ainsi l’activité transcriptionnelle de la p53 et supprimant l’expression des gènes pro-apoptotiques. De plus, l’expression ectopique de R399E-PKM2, qui forme préférentiellement une PKM2 dimérique, améliore la résistance des cellules cancéreuses du pancréas à la gemcitabine. Ces preuves suggèrent que la PKM2 contribue à l’induction de la résistance aux médicaments via un mécanisme non métabolique. Étant donné que la PKM2 dimérique agit principalement comme régulateur transcriptionnel dans les cellules cancéreuses, les activateurs de la PKM2 qui favorisent la formation de PKM2 tétramérique peuvent être efficaces pour supprimer la fonction non métabolique de la PKM2 et réduire la résistance à la gemcitabine dans les cellules cancéreuses du pancréas. Il a également été démontré que la PKM2 nucléaire contribue à la résistance de l’inhibiteur de l’EGFR dans le cancer colorectal et le cancer du poumon. Li et coll. a montré que la phosphorylation de STAT3 médiée par le PKM2 nucléaire réduit la sensibilité des cellules cancéreuses colorectales au gefitinib et la perturbation de l’interaction du PKM2 nucléaire et du STAT3 a restauré la sensibilité du gefitinib dans les cellules. Plus récemment, une autre étude a démontré que la PKM2 se transloque dans le noyau et interagit avec le poly-ADP ribose (PAR) lors de la stimulation du facteur de croissance. L’activité de liaison PAR de PKM2 est critique pour la rétention nucléaire et la transcription génique de PKM2 et est importante pour la promotion de la prolifération cellulaire et de la croissance tumorale. Ils ont conclu que l’inhibition de la fonction nucléaire PKM2 peut vaincre la résistance des cellules cancéreuses mutées par l’EGFR. Les deux études ont souligné un rôle crucial de la PKM2 nucléaire dans la médiation de la résistance aux médicaments dans les cancers et ont suggéré que le ciblage de la PKM2 nucléaire pourrait être une stratégie prometteuse pour remplacer la résistance.

La PKM2 contribue à la prolifération et à la résistance aux médicaments dans les cellules cancéreuses