a piruvát-kináz M2 a rákos sejtek több aspektusát táplálja: a celluláris anyagcserétől, a transzkripciós szabályozástól az extracelluláris jelátvitelig
biokémiai elemzés az enzimatikus aktivitás jellemzésével, amely katalizálja a laktát képződését a glükózból a sejtlizátumokban feltárta az első intracelluláris metabolikus utat, a glikolitikus utat. A glikolitikus aktivitást tartalmazó frakciók tisztításától kezdve számos úttörő kutató hozzájárult az enzimek azonosításához, amelyek az út minden lépésében részt vesznek . Ezek az eredmények építik fel modern koncepciónkat az aerob és anaerob légzés és energiatermelés cseréjében különböző fiziológiai és patológiai körülmények között.
először 1934-ben számoltak be egy enzim létezéséről, amely katalizálta az ATP termelését azáltal, hogy egy foszfátcsoportot a PEP-ből az ADP-be továbbított a májban . A piruvát-kináz (PK) néven ismert enzim későbbi izolálása eltéréseket mutatott a szöveti eloszlásban és a katalitikus kinetikában, ami arra utal, hogy ennek az enzimnek különböző izoformái lehetnek . 1982 és 1984 között különböző PK géneket klónoztak élesztőből, csirkéből és patkányból . A pkm2 funkcionális vizsgálatát egy jelölt gén azonosításával kezdeményezték egérben az 1980-as évek elején . Később, Noguchi et al. kimutatták, hogy a PK két izoformáját (PKM1 és PKM2) ugyanaz a PKM gén kódolja alternatív splicing útján . Emberben a PKM izoformákat hasonló splicing mechanizmuson keresztül állítják elő, a 9 .és 10. exont külön-külön a PKM1 és PKM2 mRNS-be integrálva.
számos megállapítás felhívta a kutató figyelmét a PKM2 lehetséges szerepére a tumorigenesisben. Először is, a PKM2 az embrionális izoforma, amely erősen expresszálódik az állatok fejlődése során. Transzkripciója számos felnőtt szövetben gyengül, miközben a daganatokban újraaktiválódik . Másodszor, a pkm1 és a pkm2 relatív bőségének vizsgálata a normál és a tumorszövetekben kimutatta a PKM1 izoformról a PKM2 izoformra való áttérést különböző rákokban, például hepatocelluláris karcinómában . Harmadszor, az mRNS splicing PKM1-ről PKM2-re történő változását fokozza a C-Myc onkogén, ami arra utal, hogy a rákos sejtek aktívan részt vesznek ebben a váltásban, hogy megfeleljenek a proliferáció és az anyagcsere követelményeinek . Negyedszer, a pkm2 aktivitásának aktivátorok vagy inhibitorok általi modulálása befolyásolja a tumor növekedését in vivo .
- az első epizód: A PKM2 mint metabolikus enzim a citoplazmában
- a második epizód: pkm2 jelátviteli modulátorként a citoplazmában
- A harmadik epizód: PKM2 mint transzkripciós szabályozó a magban
- A negyedik epizód: PKM2 mint extracelluláris jelátviteli kommunikátor
- The fifth episode: a PKM2 megválaszolatlan eltérése
- a folytatódó epizód: a PKM2 rák biomarker és gyógyszer célpont?
az első epizód: A PKM2 mint metabolikus enzim a citoplazmában
mivel a pkm2 szerepét a glikolízis metabolikus szabályozásában a rákos sejtekben alaposan áttekintették, itt csak három lényeges különbséget foglalunk össze a PKM1 és a PKM2 által közvetített katalízis és a celluláris metabolizmus között. Az első különbség az alegység interakciója. Mind a PKM1, mind a PKM2 tetramer fehérjék, amelyeket négy azonos alegység alkot. Minden alegység (vagy monomer) négy szerkezeti domént tartalmaz, beleértve az A, B, C és N-terminális domént. A monomer először dimerizálódik, majd két dimer kölcsönhatásba lép a dimer-dimer interfészen keresztül, amelyet a monomer C doménje rendez, hogy tetramert képezzen. Mivel a PKM1 és a PKM2 különböző exonokat tartalmaz az mRNS-ekben, ez megváltoztatja a kódolt aminosavakat A C doménben, és megváltoztatja a tetramer stabilitását. Fiziológiai állapotban a PKM1 konstitutívan tetramer-ként szerveződik, míg a pkm2 létezhet tetramerben vagy dimerben. A második különbség az alloszterikus szabályozás. A kis molekulák és metabolitok intracelluláris koncentrációjától függően a PKM1 és a PKM2 aktivitása differenciáltan szabályozható. Az egyik legismertebb alloszterikus szabályozó a fruktóz-1,6-biszfoszfát (FBP). Ez a glikolitikus köztitermék közvetlenül kötődik a PKM2-hez, és növeli a PKM2 affinitását a PEP-hez . Éppen ellenkezőleg, az FBP nem befolyásolja jelentősen a PKM1 aktivitását. Az FBP mellett más metabolitokról, aminosavakról és kis molekulákról is beszámoltak, amelyek befolyásolják a PKM2 aktivitást (ábra. 1). Az aktiváláshoz vagy gátláshoz szükséges koncentráció azonban magas, a moduláló hatás pedig mérsékelt. Még nem tisztázott, hogy ezek a kis molekulák fontos szerepet játszanak-e a PKM aktivitásának szabályozásában fiziológiai körülmények között. A PKM2 aktivitást a poszttranszlációs módosítás is szabályozza, mint például a foszforilezés, az acetilezés és az oxidáció, amelyek elősegítik a dimer PKM2 alacsony aktivitását (ábra. 1). A harmadik különbség az energiatermelés és a köztes felhasználás. Mivel a PKM1 konstitutívan aktív tetramer, ennek az izoformának a fő biológiai funkciója az ATP előállítása a sejtenergia ellátására. A PKM2 azonban az ATP előállítása mellett átállhat a kevésbé aktív dimer formára, hogy több glikolitikus intermediert hozzon létre, amelyek építőelemként használhatók az aminosavak, lipidek és nukleotidok bioszintéziséhez.
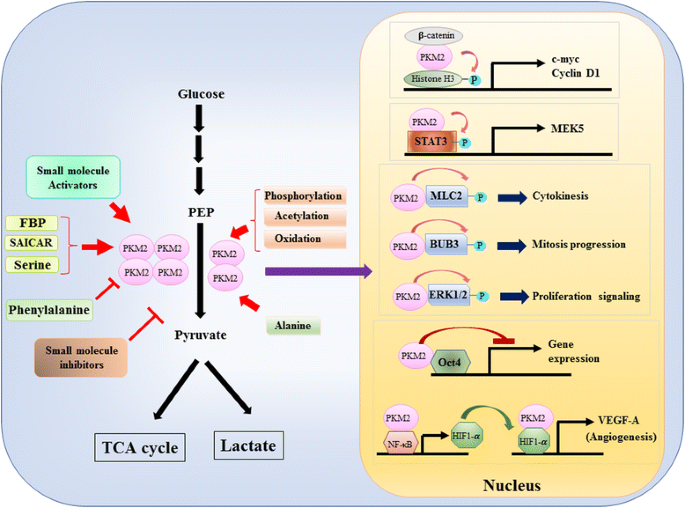
a pkm2 aktivitás modulációja fiziológiai molekulákkal és poszttranszlációs módosítással, valamint a PKM2 potenciális szerepe a transzkripciós szabályozásban
a második epizód: pkm2 jelátviteli modulátorként a citoplazmában
a glikolitikus enzimként való működés mellett a pkm2-t javasolják több sejtes folyamatba bevonni a citoplazmában kölcsönhatásba lépő fehérjék azonosítása miatt. Például kimutatták, hogy a PKM2 több tirozin-kináz kölcsönhatásba lépő fehérje, beleértve az A-Raf-ot, a töréspont klaszter régió-Abelson (BCR-ABL) fúziós kináz, fibroblaszt növekedési faktor receptor 1 (FGFR1) stb. . Kimutatták, hogy ezek a kötő partnerek modulálják a pkm2 dimer/tetramer változását a sejtek anyagcseréjének megváltoztatására. Lehetséges azonban, hogy a pkm2 kölcsönösen befolyásolhatja ezen kötő partnerek katalitikus kinetikáját, szubsztrátkötését és citoplazmatikus elhelyezkedését a jelátvitel modulálása érdekében. Az a megállapítás, hogy a PKM2 foszfor-tirozin kötő fehérje, megerősíti ezt a lehetőséget, mivel sok intracelluláris jelátviteli mediátor kötődhet a foszfor-tirozin maradékhoz, hogy specifikus fehérjekomplexeket állítson össze a jelátvitelhez . A mai napig a pkm2 kötő partnerek listája folyamatosan növekszik. Számos új tagot emelünk ki, és itt tárgyaljuk biológiai vonatkozásaikat. Mukheriee et al. kimutatták, hogy a PKM2 képes kötődni a HuR-hoz, egy RNS-kötő fehérjéhez, amely fontos szerepet játszik az mRNS stabilitásának és transzlációs hatékonyságának szabályozásában, elősegítve a sejtciklus progresszióját és a glioma sejtek proliferációját . Érdekes módon egy másik RNS-kötő fehérje, a tristetraprolin, amely számos mRNS-t köthet az AU-gazdag elemen keresztül a 3 ‘- nem transzlációs régióban (3 ‘ – UTR), szintén pkm2 kölcsönhatásban lévő partnernek bizonyult, és a PKM2 indukálta a tristetraprolin foszforilációját és lebomlását az emlőrák növekedésének modulálása érdekében . Ez a két tanulmány a PKM2 potenciális transzlációs kontroll funkcióját jelenti. Nemrég, Liang et al. a bcl2 anti-apoptotikus fehérjét új PKM2 partnerként azonosította . Kimutatták, hogy az oxidatív stressz indukálta a pkm2 transzlokációját mitokondriumokba, ahol foszforilálta és stabilizálta a Bcl2 – t azáltal, hogy megakadályozta annak lebontását ubiquitináció-függő úton. Ezek az adatok arra utaltak, hogy a PKM2 segíti a rákos sejteket az intracelluláris metabolikus változás vagy extracelluláris sértés által kiváltott oxidatív stressz adaptálásában.
A harmadik epizód: PKM2 mint transzkripciós szabályozó a magban
a pkm2 nukleáris szerepét a gén transzkripciójának vagy epigenetikus módosításának szabályozásában először az a megállapítás javasolta, hogy a PKM2 kötődik Y333-foszforral foszforral rendelkező caeton-kateninhez, és a caeton-katenin-PKM2 komplexet toborozták a nukleoszómákba, hogy foszforilálják a H3 hisztont a treonin 11-nél (ábra. 1). Ez a foszforilezés ezt követően fokozta a hiszton H3 acetilezését, ami az a-katenin célgének szabályozása. A PKM2 által közvetlenül foszforilált másik transzkripciós faktor a jelátalakító és a 3 transzkripció aktivátora (STAT3) . A STAT3 pkm2 által közvetített foszforilezése a tirozin 705-nél fokozta a STAT3 aktivitást a mitogén-aktivált protein-kináz kináz 5 (MEK5) expressziójának szabályozására. A transzkripciós faktorok mellett kimutatták, hogy a PKM2 foszforilálja a miozin 2-es könnyű láncot (MLC2), a BUB3-at és az extracelluláris szignál-szabályozott kináz 1-et és 2-t (ERK1 és ERK2) . Érdekes módon a PKM2 foszforilációtól független módon is hat a génexpresszió befolyásolására. Például a PKM2 kötődik az Oct4-hez, az egyik mester transzkripciós faktorhoz, amely szabályozza az őssejtek önmegújulását, és gátolja az Oct4 által közvetített transzkripciót . A PKM2 fokozhatja a tumor angiogenezisét azáltal, hogy kölcsönhatásba lép az NF-kB-val és a HIF-1-vel a sejtmagban, és aktiválja a HIF-1-et a célgén VEGF-A expresszióját .következésképpen a VEGF-A fokozott szekréciója fokozza az erek képződését, ami hozzájárul a tumor növekedéséhez. Bár ezek a vizsgálatok erősen sugallták a pkm2 nukleáris lokalizációját és protein kináz funkcióját különböző fiziológiai és patológiai körülmények között, a nukleáris PKM2 által közvetített génexpresszió fontosságát azonban a pkm2 knockout sejtekkel végzett vizsgálatok megkérdőjelezték. A-jelölt PEP és PKM2-null egér embrionális fibroblasztok használatával Hosios et al. kimutatták, hogy a PEP-függő foszforiláció nem gyakori esemény a sejtekben, és a reakciót nem katalizálja a PKM2 . Ezeknek a vizsgálatoknak az eltérése jelenleg megoldatlan, és a pkm2 protein-kináz aktivitása további megerősítést igényel.
A negyedik epizód: PKM2 mint extracelluláris jelátviteli kommunikátor
az extracelluláris PKM2 jelenléte új utat nyit a pkm2 biológiai funkciójának tanulmányozásához. Buschow et al. az első bizonyítékot szolgáltatta arra, hogy a PKM2 kimutatható a B-sejt exoszómákban, és MHC II .osztályba tartozó fehérjeként azonosították. Két későbbi tanulmány azt is jelezte, hogy a PKM2 létezik a különféle sejtek által felszabadított exoszómákban . Jelenleg számos nyilvános adatbázis, mint például az ExoCarta és az EVpedia átfogó információt nyújt a különböző fajok extracelluláris vezikulumainak fehérjéiről, lipidjeiről, nukleinsavairól. Az összes adat megerősíti, hogy a PKM2 az exoszómák csomagfehérje. A legújabb tanulmányok egyértelműen kimutatták az exoszómák kommunikációs szerepét azáltal, hogy különböző komponenseket szállítanak a gazdasejtekből a recipiens sejtekbe . Várható, hogy a PKM2 szerepet játszhat a sejt-sejt áthallásban.
a feltörekvő bizonyítékok valóban alátámasztják ezt a hipotézist. Például egy nemrégiben készült tanulmány kimutatta, hogy a vérben keringő PKM2 elősegítheti a tumor növekedését és az angiogenezist azáltal, hogy növeli az endothel sejtek növekedését, migrációját és mátrix adhézióját . Egy másik vizsgálat azt is kimutatta, hogy a vastagbélrákos sejtekből szekretált PKM2 autokrin stimuláció révén fokozhatja a sejtvándorlást a PI3K/Akt és a Wnt/MHz-catenin útvonalak aktiválásával . A rákos sejtek mellett a szövetkárosodás helyén lévő neutrofilek felszabadíthatják a PKM2-t az angiogenezis és a sebgyógyulás elősegítése érdekében . Nemrégiben végzett Vizsgálatunk azt is kimutatta, hogy a rekombináns PKM2 fehérje indukálhatja az epidermális növekedési faktor receptor (EGFR) foszforilációját és aktiválódását . Sőt, azt találtuk, hogy r339e mutáns PKM2 amely előnyösen kialakított dimer PKM2 aktivált EGFR szignifikánsan nagyobb, mint a tetramer PKM2. Keller és munkatársai. a Hela sejtek szukcinil-5-aminoimidazol-4-karboxamid-1-ribóz-5′-foszfáttal (SAICAR) történő kezelését követően 154 fehérjét azonosítottak a Pkm2 protein-kináz aktivitását serkentő intracelluláris metabolitként . Az EGFR-t PKM2 szubsztrátként is megtalálták. Eredményeik két szempontból különböznek a miénktől. Először is, a vizsgálatunkban aktivált jelátviteli utakat extracelluláris PKM2 váltja ki, míg a vizsgálatukban azonosított molekuláris célpontok az intracelluláris PKM2 potenciális szubsztrátjai. Másodszor, az ERK1/2 aktivitás növekedését vizsgálatunkban EGFR aktiválás kezdeményezi, míg az ERK1/2 aktivációt tanulmányukban közvetlenül stimulálja a SAICAR/PKM2 komplex. Az egyik hasonló jelenség, amelyet mindkét vizsgálatban megfigyeltek, az, hogy az R339E mutáns PKM2 szignifikánsabban aktiválja a jelátviteli molekulákat, mint a vad típusú PKM2, ami arra utal, hogy a dimer és a tetramer PKM2 külön szerepet játszik az onkogenezisben. A receptor tirozin-kináz tömb alkalmazásával azt találtuk, hogy az extracelluláris PKM2 csak korlátozott növekedési faktor receptorokat aktivált az emlőrákos sejtekben (az adatok nem jelennek meg). Jelenleg az extracelluláris pkm2 receptor aktiválásának szelektivitása továbbra sem ismert. Ezenkívül nem világos, hogy az R339E mutáns PKM2 miért hatékonyabb az EGFR aktiválásában. További kísérletekre van szükség e kérdések megválaszolásához.
egy másik elegáns kérdés, amellyel foglalkozni kell, hogy a szabad PKM2 és a vezikulacsomagolás pkm2 hasonló hatást fejtenek-e ki a tumorigenesis elősegítésében (ábra. 2). A szabad extracelluláris PKM2 nem tudott behatolni a plazmamembránba, és csak sejtfelszíni fehérjéken, például növekedési faktor receptorokon keresztül tudta aktiválni az intracelluláris jelátvitelt. Ezzel szemben a vezikulumcsomagoló PKM2-t rákos és sztrómasejtek endocitizálják, és a vezikulákból felszabaduló PKM2 intracelluláris mechanizmuson keresztül befolyásolhatja az anyagcserét és a génexpressziót. The PKM2-null cells or mice will be useful to elucidate whether these two extracellular forms of PKM2 could act synergistically or antagonistically in tumorigenesis.

The potential tumor-promoting effect of extracellular PKM2
The fifth episode: a PKM2 megválaszolatlan eltérése
a fent tárgyalt sejtalapú adatok mellett a pkm2 onkogén szerepét is megkérdőjelezték a pkm2 kiütéses egerek generálása után. Israelsen et al. feltételes knockout egér modellt generált a PKM2-specifikus 10. exon törlésével . Meglepő módon a pkm2 kimerülése felgyorsította, de nem gyengítette a tumorképződést, amelyet a BRCA1 gén elvesztése vezetett egerekben. Ezek az adatok azt mutatták, hogy a pkm2 nem szükséges a rákos sejtek proliferációjához. Érdekes módon a PKM1 expressziót csak a nem proliferáló tumorsejtekben észlelték, ami a pkm1 tumorszuppresszív szerepére utal az emlőrákban. Ezenkívül a PKM2 knockout egereknek magas az előfordulási gyakorisága a hepatocelluláris karcinóma spontán kialakulása hosszú késleltetés után az anyagcsere egyensúlyhiánya miatt . Ezek az eredmények azzal a gondolattal szemben, hogy a PKM2 in vivo onkogén szerepet játszik.
a folytatódó epizód: a PKM2 rák biomarker és gyógyszer célpont?
bár a géntechnológiával módosított egérmodell eredményei nem támasztják alá a pkm2 tumort elősegítő aktivitását, a pkm2 túlzott expressziója általánosan megtalálható az emberi daganatokban, és rossz klinikai eredménnyel jár (1 .táblázat). Két újabb metaanalízis tanulmány is alátámasztotta ezt a következtetést. Wu et al. elemezte az adatokat a 2812 betegek szilárd tumorok emésztőrendszer nyert 16 kohorsz vizsgálatok és megállapította, hogy a túlzott expressziója PKM2 társul csökkent teljes túlélés gyomorrák, nyelőcső laphámsejtes karcinóma, hepatocelluláris karcinóma, epeúti rák és a szájüregi rák . A PKM2 azonban nem prognosztikai tényező a hasnyálmirigyrákban. Ez a megállapítás nem áll összhangban a korábbi tanulmányokkal, amelyek azt mutatják, hogy a PKM2 növekedése a hasnyálmirigyrákos betegek rossz túlélésének mutatója . Egy másik vizsgálat 4796 esetet tartalmazott 27 egyedi vizsgálatból, amelyek kimutatták, hogy a PKM2-emelkedés összefügg a rosszabb teljes túléléssel, a betegségmentes túléléssel és a kiújulás nélküli túléléssel az összesített adatokban . Ráktípus szerint rétegezve azonban a PKM2 nem jósolja meg a hasnyálmirigyrák rossz túlélését. Összességében úgy tűnik, hogy a pkm2 felelős prognosztikai marker a legtöbb szilárd daganatban.
éppen ellenkezőleg, a pkm2 diagnosztikai tényezőként történő használata ellentmondásos. Egy proteomikus elemzés kimutatta, hogy a PKM2 potenciális diagnosztikai marker a tüdőrák kimutatására . Egy nemrégiben készült tanulmány szerint azonban a PKM2 nem jó diagnosztikai marker a tüdőrákban az alacsony specifitás miatt . Hasonlóképpen, a PKM2 önmagában valószínűleg nem lesz hasznos marker a vastagbélrák szűrésében . Azonban több marker kombinációja növelheti a rák diagnózisának érzékenységét és specifitását .
a pkm2 terápiás potenciálja érdekes esemény a rák kezelésében. Az egyik oldalról a PKM2 gátlása várhatóan gátolja a glikolízist, rontja a gén transzkripciót és elnyomja a sejtproliferációt. Ezért úgy tűnik, hogy a PKM2 inhibitorok jó jelöltek a rákellenes gyógyszerek fejlesztésére. A könyvtári szűrés használatával, Vander Heiden et al. a PKM2 inhibitorok három új osztályát azonosította, és kimutatta, hogy a leghatékonyabb vegyület gátolta a pkm2 aktivitást és indukálta a rákos sejtek halálát . Nemrég, Ning et al. megállapította, hogy az új naftokinon-származékok erős PKM2 inhibitorok . Egy hatékony vegyület 3 k elnyomta több rákos sejtvonal proliferációját szubmikromoláris koncentrációban, miközben kevés káros hatást mutatott a normál sejtekre. A másik oldalról a PKM2 aktiválása szintén gátolhatja a tumor növekedését. Mivel az alacsony aktivitású PKM2 dimer a fő izoforma, amely kiváltja a glikolízist a citoplazmában és a gén transzkripcióját a sejtmagban a rákos sejtekben, a pkm2 aktivátorok, amelyek elősegíthetik a tetramer PKM2 képződését, a glikolízist mitokondrium útvonalra válthatják, és csökkenthetik a nukleáris belépést, hogy gyengítsék a gén transzkripcióját. Mindkét hatás rontja az anyagcsere-keresletet és a növekedést támogató jelátvitelt, amely tumor regresszióhoz vezet. Két úttörő vizsgálatban különböző PKM2 aktivátorokat azonosítottak, és in vitro jellemezték specificitásukat . Egy későbbi tanulmány kimutatta, hogy a PKM2 aktivátorok valóban elősegítették a tetramer képződését és elnyomták a tumor növekedését in vivo . Ezek az eredmények azt sugallták, hogy a PKM2 aktivátorok ígéretes rákellenes gyógyszerek lehetnek.
a kemoterápiával szembeni rezisztencia a rák kezelésének egyik fő elzáródása. A rákos sejtek inherens kemorezisztenciájának leküzdése sürgős a klinikai kutatáshoz. Kimutatták a pkm2 szerepét a rákos sejtek kemorezisztenciájában, és kimutatták, hogy a PKM2 célzása újra szenzibilizálja a kemorezisztens rákos sejteket. Egy nemrégiben készült tanulmány kimutatta, hogy a CD44 kölcsönhatásba lép a PKM2-vel, és elnyomja a PKM2 aktivitását a Pkm2 tyr105 foszforilációjának növelésével. A CD44 abláció indukálta az aerob glikolízisről a mitokondriális légzésre való áttérést és a reaktív oxigénfajok (ROS) termelésének fokozását, ami a kolorektális rákos sejtek ciszplatin érzékenységének fokozódását eredményezte . Kimutatták, hogy a pkm2 aktivitás gátlása gátolja a glikolízist és legyőzi a ciszplatin rezisztenciát. Ezért a ciszplatin és a PKM2 inhibitorok kombinációja hatékony stratégia lehet a kemoterápiában (ábra. 3). Beszámoltak arról is, hogy a pkm2 részt vesz a gemcitabin-rezisztencia szabályozásában hasnyálmirigyrákos sejtekben. Kim et al. kimutatták, hogy a PKM2 elősegíti a gemcitabin rezisztenciát a p38 által közvetített p53 foszforiláció negatív szabályozásával, ezáltal csökkentve a p53 transzkripciós aktivitását és elnyomva a pro-apoptotikus gének expresszióját . Ezenkívül az r399e-PKM2 méhen kívüli expressziója, amely előnyösen dimer PKM2-t képez, fokozza a hasnyálmirigyrák sejtek rezisztenciáját a gemcitabinnal szemben. Ezek a bizonyítékok arra utalnak, hogy a PKM2 nem metabolikus mechanizmuson keresztül járul hozzá a gyógyszerrezisztencia kiváltásához. Mivel a dimer PKM2 főként transzkripciós szabályozóként működik a rákos sejtekben, a pkm2 aktivátorok, amelyek elősegítik a tetramer PKM2 képződését, hatékonyan gátolhatják a pkm2 nem metabolikus funkcióját és csökkenthetik a gemcitabinnal szembeni rezisztenciát a hasnyálmirigyrákos sejtekben. Kimutatták, hogy a nukleáris PKM2 hozzájárul az EGFR inhibitor rezisztenciájához a vastagbélrákban és a tüdőrákban . Li et al. kimutatták, hogy a nukleáris PKM2-mediált STAT3 foszforiláció csökkenti a colorectalis rákos sejtek érzékenységét a gefitinibbel szemben, és megszakítja a nukleáris PKM2 és STAT3 helyreállított gefitinib érzékenység kölcsönhatását a sejtekben . A közelmúltban egy másik tanulmány kimutatta, hogy a PKM2 transzlokálódik a magba, és kölcsönhatásba lép a poli-ADP ribózzal (par) a növekedési faktor stimuláció során. A PKM2 PARKÖTŐ aktivitása kritikus a pkm2 nukleáris retenciója és géntranszkripciója szempontjából, és fontos a sejtproliferáció és a tumor növekedésének elősegítése szempontjából. Arra a következtetésre jutottak, hogy a pkm2 nukleáris funkció gátlása leküzdheti az EGFR-mutált rákos sejtek rezisztenciáját . Mindkét tanulmány rámutatott a nukleáris PKM2 kulcsfontosságú szerepére a rákos megbetegedések gyógyszerrezisztenciájának közvetítésében, és azt javasolta, hogy a nukleáris PKM2 megcélzása ígéretes stratégia lehet a rezisztencia felülírására.

a PKM2 hozzájárul a rákos sejtek proliferációjához és gyógyszerrezisztenciájához