C-Vitamin: antioxidáns szer
A C-vitamin redox metabolizmusa és antioxidáns tulajdonságai
a szabad gyökök és oxidánsok kettős szerepet játszanak mind mérgező, mind hasznos vegyületekként az anyagcsere folyamatokban és az exogén stimulációkra adott válaszként. Ezek vagy normális anyagcsere-tevékenységekből, vagy környezeti tényezőkből (szennyezés, cigarettafüst, sugárzás) származnak. Ha a szabad gyökök túlterhelését nem lehet megsemmisíteni, a testben történő felhalmozódásuk oxidatív stresszt generál . Az oxidatív stressz akkor fordul elő, amikor a szabad gyökök képződése meghaladja az ellenük való védelem képességét. Ez a folyamat vezet a krónikus és degeneratív betegségek, mint a rák, autoimmun betegségek, öregedés, szürkehályog, rheumatoid arthritis, szív-és érrendszeri, és neurodegeneratív betegségek . Az antioxidáns olyan molekula, amely megakadályozza más molekulák oxidációját. Az oxidációs folyamat olyan kémiai reakció, amely szabad gyököket termel, ami láncreakciókhoz vezet, amelyek károsíthatják a sejteket. A C-vitamin antioxidáns hatását jól dokumentálták . A C-Vitamin egy erős antioxidáns, amely képes hidrogénatomot adományozni, és viszonylag stabil aszkorbil – szabad gyököt alkot. Az E-Vitamin, A C-vitamin és a karotin antioxidáns vitaminokként ismertek, amelyek csökkentik az oxidatív károsodást és csökkentik bizonyos krónikus betegségek kockázatát. A betegségek, mint például a kardiovaszkuláris rendellenességek, az epidemiológiai vizsgálatok során az l-aszkorbinsav, a tokoferol és a Mhz-karotin nem megfelelő koncentrációjával járnak. A C-Vitamin fokozza a vas felszívódását azáltal, hogy a Fe3+-t Fe2+ – ra csökkenti a nem hem vasforrásokból . Redox-aktív ionok (vas, réz) jelenlétében a C-vitamin prooxidánsként működik, hozzájárulva a hidroxilgyökök képződéséhez, ami lipidhez vezethet, DNS, vagy fehérje oxidáció . Különböző mechanizmusok léteznek az oxidatív stressz enyhítésére és a sérült makromolekulák helyreállítására. Az enzimatikus és nem enzimatikus antioxidánsok fontos szerepet játszanak a szabad gyökök és a reaktív oxigénfajok (ROS) eltávolításában. Kimutatták, hogy az oxidatív stressz jelentősen befolyásolja az antioxidáns enzimeket, a katalázt (CAT), a szuperoxid-diszmutázt (sod), a glutation-reduktázt (GR), a glutation-peroxidázt (gshpx) és a növényekben az aszkorbát-peroxidázt (AA-px) és a nemenzimes antioxidánsokat, beleértve a glutationt (GSH) és az aszkorbátot (ASC). Az antioxidáns vegyületek megakadályozhatják a szabad gyökök ellenőrizetlen képződését, vagy gátolhatják a biológiai helyekkel való reakciójukat; továbbá a legtöbb szabad gyök pusztulása az endogén antioxidánsok oxidációjától függ, elsősorban a molekulák eltávolításával és redukálásával . Úgy gondolják, hogy a C-Vitamin fontos vízben oldódó antioxidáns, amely a jelentések szerint semlegesíti a ROS-t és csökkenti az oxidatív stresszt .
A C-Vitamin hatékony redukálószer és a szabad gyökök elpusztítója a biológiai rendszerekben . Részt vesz az antioxidáns védelem első sorában, védi a lipidmembránokat és a fehérjéket az oxidatív károsodástól. Vízben oldódó molekulaként a C-vitamin a sejteken belül és kívül is működhet, semlegesítheti a szabad gyököket és megakadályozhatja a szabad gyökök károsodását. A C-Vitamin kiváló elektronforrás a szabad gyökök számára, amelyek egy elektront keresnek, hogy visszanyerjék stabilitásukat. A C-Vitamin elektronokat adhat a szabad gyököknek, és leállíthatja reakcióképességüket .
A C-vitaminról kimutatták, hogy hatékony tisztítószer az oxigén és a nitrogén-oxid fajok ellen, mint például a szuperoxid gyökös ion, a hidrogén-peroxid, a hidroxilgyök és a szingulett oxigén. A C-vitamin ezen tulajdonsága létfontosságú folyamatokkal rendelkezik a sejtkomponensek védelmében a szabad gyökök által kiváltott károsodásoktól. Ezenkívül a C-vitamin hatékonyan regenerálja az E-vitamin antioxidáns formáját a tokoferoxilgyökök csökkentésével. Ez a folyamat megvédi a membránokat és a sejt egyéb rekeszeit a szabad gyökök által kiváltott károsodástól (2.ábra). Az aszkorbát-peroxidáz (APX) egy enzim, amely redukálja a H-T2O2 vízbe az aszkorbát elektrondonorként történő felhasználásával. A monodehidroaszkorbát egy oxidált aszkorbát, amelyet a monodehidroaszkorbát reduktáz (MDAR) regenerál. A monodehidroaszkorbát gyök gyorsan aránytalanul aszkorbáttá és dehidroaszkorbáttá alakul. Dehydroascorbate is reduced to ascorbate by dehydroascorbate reductase in the presence of GSH, yielding oxidized glutathione (GSSG). It is reduced by glutathione reductase (GR) using nicotinamide adenine dinucleotide phosphate hydrogen (NADPH) as an electron donor. Dehydroascorbate may be reduced nonenzymatically or catalyzed by proteins with dehydroascorbate reductase (DHAR) activity.
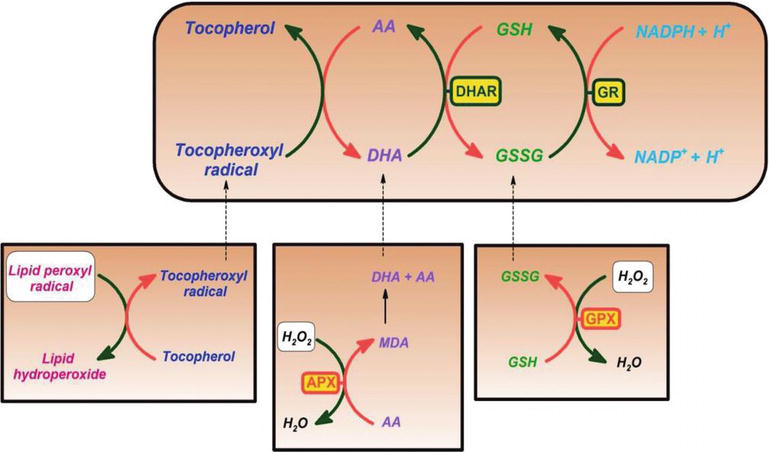
Figure 2.
xmlns:mml=”http://www.w3.org/1998/Math/MathML” xmlns:xlink=”http://www.w3.org/1999/xlink” xmlns:xsi=”http://www.w3.org/2001/XMLSchema-instanceAscorbate and redox cycling antioxidants. AA, ascorbate; DHA, dehydroascorbate; DHAR, semidehydroascorbate reductase; GSH, glutathione; GSSG, semi-glutathione reductase; GR, glutathione reductase; APX, ascorbate peroxidase; and GPX, glutathione peroxidase .
Glutathione-ascorbate cycle operates in the cytosol, mitochondria, plastids, and peroxisomes in plants . Javasolt, hogy a glutation-aszkorbát ciklus kulcsszerepet játszik a H2O2 méregtelenítésében, mivel a glutation, az aszkorbát és a NADPH magas koncentrációban van a növényi sejtekben. Más enzimek, mint például az aszkorbát és a glutation-peroxidázok, amelyek tioredoxinokat vagy glutaredoxinokat használnak redukáló szubsztrátként, szintén szerepet játszanak a H2O2 eltávolításában a növényekben (2.ábra).
A C-Vitamin szintén képezi a szemidehidroaszkorbil gyököt, egy viszonylag hosszú élettartamú gyököt, az E-vitamin regenerálásában radikális formájából, valamint a gyökök eltávolításában. A növényi és állati sejtek NADH-függő szemidehidroaszkorbát-reduktáz enzimet (EC 1.6.5.4) tartalmaznak, amely redukálószerként NADH-t használva a gyököt C-vitaminná redukálja (2.ábra). Mind enzimatikusan, mind nemenzimatikusan visszafordíthatatlanul bomlik diketoglükonsavvá, vagy glutationfüggő reakcióban aszkorbáttá alakítható .
redukáló anyagként és elektrondonorként a szabad gyökök eltávolítása során a C-vitamin nagy energiájú elektronokat adományoz a szabad gyökök semlegesítésére, és dehidroaszkorbinsavvá oxidálódik. A dehidroaszkorbinsav újra felhasználható aszkorbinsavvá, vagy metabolizálható, további elektronok felszabadításával. Bár a C-vitamin a bélből egy nátriumfüggő C-vitamin transzporteren keresztül szívódik fel, a legtöbb sejt oxidált formában (dehidroaszkorbinsav) szállítja a C-vitamint glükóz transzporteren keresztül 1. A dehidroaszkorbinsav redukálódik, hogy aszkorbinsavat hozzon létre a sejt belsejében, megvédve a mitokondriumokat a szabad gyökök által kiváltott oxidatív károsodástól (2.és 3. ábra). Erősen reaktív szabad gyökök (pl., RO–, RO2–, OH -,, NO2) aszkorbáttal redukálódnak, az újonnan keletkezett aszkorbilgyök pedig rosszul reagálnak. Az aszkorbát a peroxinitrit, például hipoklórsavból, ózonból és nitráló szerekből származó nem radikális reaktív fajokat is elpusztíthat. A C-Vitamin egy monoszacharid oxidációcsökkentő (redox) katalizátor, amely mind állatokban, mind növényekben megtalálható. A C-vitamin antioxidáns hatása annak köszönhető, hogy képes elektronokat adományozni mind a második, mind a harmadik szénből. A főemlősök evolúciója során az aszkorbinsav előállításához szükséges enzimek egyike elveszett a mutáció miatt, az embereknek az étrendből kell beszerezniük ; a legtöbb állat képes szintetizálni ezt a vitamint a testükben, és nem szükséges az étrendjükben . C-vitaminra van szükség a prokollagén kollagénné történő átalakításához azáltal, hogy a prolinmaradványokat hidroxiprolinná oxidálja. Más sejtekben redukált formában tartják fenn a glutationnal való reakcióval . Amint azt a 2.és 3. ábra mutatja, az aszkorbinsav redox katalizátor, amely képes csökkenteni és ezáltal semlegesíteni a ROS-t, például a hidrogén-peroxidot (H2O2) (2. és 3. ábra).
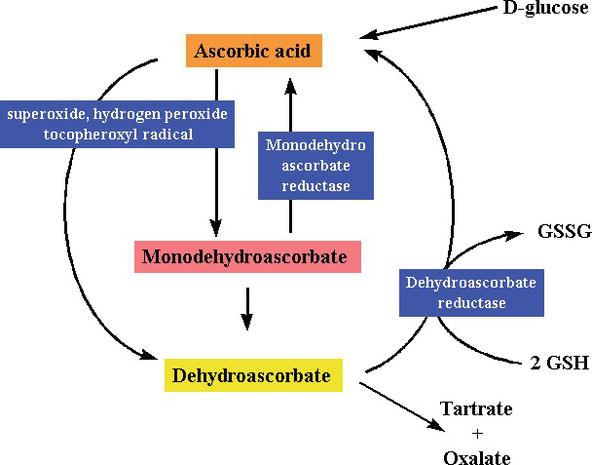
Figure 3.
xmlns:mml=”http://www.w3.org/1998/Math/MathML” xmlns:xlink=”http://www.w3.org/1999/xlink” xmlns:xsi=”http://www.w3.org/2001/XMLSchema-instanceSynthesis and degradation ofl-ascorbic acid in plant tissues .
Ascorbic acid has direct antioxidant effects, and also it is a substrate for the redox enzyme ascorbate peroxidase, that is particularly important in stress resistance in plants. Az aszkorbinsav magas szinten van jelen a növények minden részében, különösen azokban a kloroplasztokban, amelyek ott 20 mM-es koncentrációt érnek el . Dehidroaszkorbát (DHA) és aszkorbát szabadgyök (AFR), mint köztitermék, az aszkorbát szabadgyök (AFR), amelyek reverzibilis, egyelektronos oxidációk keletkeznek aszkorbátból (4.ábra). A Ros enzimatikus eltávolításának általánosan feltételezett modellje szerint a SOD katalizálja a szuperoxid aniont H2O2-vé és oxigénné; majd a H2O2-t vízre és molekuláris oxigénre redukálja a CAT. CAT turnover number is very high, but its affinity for H2O2 is relatively low, and consequently a certain amount of H2O2 remains in the cell.
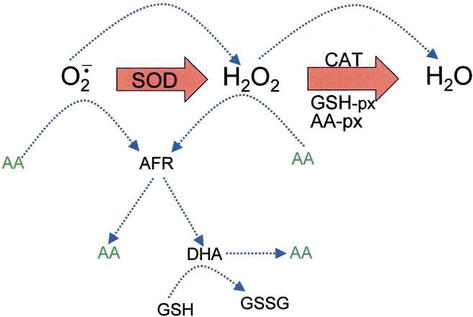
Figure 4.
xmlns:mml=”http://www.w3.org/1998/Math/MathML” xmlns:xlink=”http://www.w3.org/1999/xlink” xmlns:xsi=”http://www.w3.org/2001/XMLSchema-instanceThe role of AA in the detoxification of ROS. Blue dotted lines indicate nonenzymatic reactions.
H2O2 can react with superoxide anion formed in oxidative metabolism generating the highly reactive hydroxyl radical. GSH peroxidases (GSH-px) and AA peroxidases (AA-px) are capable of scavenging H2O2 due to their high affinity for H2O2. The cooperativity of SOD, CAT, and peroxidases ensures low amounts of superoxide anion and H2O2 and limiting the risk of hydroxyl radical formation (Figure 5).

Figure 5.
xmlns:mml=”http://www.w3.org/1998/Math/MathML” xmlns:xlink=”http://www.w3.org/1999/xlink” xmlns:xsi=”http://www.w3.org/2001/XMLSchema-instanceFoyer-Halliwell-Asada cycle .