ピルビン酸キナーゼM2燃料癌細胞の複数の側面:細胞代謝から、転写調節から細胞外シグナル伝達へ
細胞溶解物中のグルコースから乳酸の形成を触媒する酵素活性を特徴付けることによる生化学的分析は、最初の細胞内代謝経路、解糖経路を明らかにした。 解糖活性を含む画分の精製から始めて、多くの先駆的研究者が経路の各ステップに関与する酵素の同定に貢献した。 これらの結果は、様々な生理学的および病理学的状況下での好気性および嫌気性呼吸およびエネルギー生産の交換における現代的な概念を構築する。
肝臓でpepからADPにリン酸基を移動させることによってATPの産生を触媒する酵素の存在が最初に報告されたのは1934年である。 その後のピルビン酸キナーゼ(PK)として知られる酵素の単離は、この酵素が異なるアイソフォームを有する可能性があることを示唆している組織分布と触媒速度論の違いを示した。 1982年から1984年にかけて、酵母、ニワトリ、ラットから様々なPK遺伝子がクローニングされた。 PKM2の機能研究は、1980年代初頭にマウスの候補遺伝子の同定によって開始されました。 後に、野口らは、””を発表している。 PKの二つのアイソフォーム(PKM1とPKM2)は、代替スプライシングを介して同じPKM遺伝子によってコードされていることを示した。 ヒトでは、PKMアイソフォームは、エクソン9と10をPKM1とPKM2mRNAに別々に含めることによって、同様のスプライシング機構を介して生成されます。
いくつかの知見は、腫瘍形成におけるPKM2の潜在的な役割に研究者の注目を集めました。 第一に、PKM2は、動物の発達中に高度に発現する胚性アイソフォームである。 その転写は、多くの成体組織において弱毒化され、一方、腫瘍において再活性化される。 第二に、正常および腫瘍組織におけるPKM1およびPKM2の相対的な存在量の研究は、肝細胞癌のような様々な癌におけるPKM1アイソフォームからPKM2イソ 第三に、PKM1からPKM2へのmRNAスプライシングの変化は、癌細胞が積極的に増殖と代謝における彼らの要件に合わせて、このスイッチに従事することを 第四に、活性化剤または阻害剤によるPKM2活性の調節は、in vivoでの腫瘍増殖に影響を及ぼす。
最初のエピソード: 細胞質における代謝酵素としてのPKM2
癌細胞における解糖の代謝制御におけるPKM2の役割が広く検討されているので、我々はPKM1とPKM2を介した触媒作用と細胞代謝の間の三つの重要な違いをここに要約するだけである。 最初の違いはサブユニットの相互作用です。 PKM1とPKM2は、四つの同一のサブユニットによって形成された四量体タンパク質である。 各サブユニット(または単量体)は、A、B、C、およびN末端ドメインを含む四つの構造ドメインを含む。 モノマーは最初に一緒に二量体化し、次に二つの二量体はモノマーのCドメインによってオーケストレーションされた二量体-二量体界面を介して相互作用して四量体を形成する。 PKM1およびPKM2がmrnaに異なったエクソンを含んでいるので、これはCの範囲の符号化されたアミノ酸を変え、四量体の安定性を変える。 生理学的条件下では、PKM1は四量体として構成的に組織化され、PKM2は四量体または二量体に存在することができる。 第二の違いは、アロステリック規制です。 小分子および代謝産物の細胞内濃度に応じて、PKM1およびPKM2の活性を差動的に調節することができる。 最もよく知られているアロステリックレギュレータの一つは、フルクトース-1,6-ビスリン酸(FBP)です。 この解糖中間体はPKM2に直接結合し、PEPに対するPKM2の親和性を増加させる。 逆に、FBPはPKM1活性に有意に影響しない。 FBPに加えて、他の代謝産物、アミノ酸および小分子が、PKM2活性に影響を及ぼすことが報告されている(図1 0A)。 1). しかし、活性化または阻害に必要な濃度は高く、調節効果は控えめである。 これらの小分子が生理的状況下でPKM活性の制御に重要な役割を果たすかどうかはまだ不明である。 PKM2活性は、リン酸化、アセチル化および酸化のような翻訳後修飾によっても調節され、これは二量体PKM2の低い活性を支持する(図1 0A)。 1). 第三の違いは、エネルギー生産と中間利用です。 PKM1は活性四量体として構成的に存在するので、このアイソフォームの主な生物学的機能は、細胞エネルギーを供給するATPの生成である。 しかし、PKM2は、ATPを産生することに加えて、活性の低い二量体形態に切り替えることができ、アミノ酸、脂質およびヌクレオチドの生合成のためのビルディングブロックとして使用することができるいくつかの解糖中間体を生成することができる。
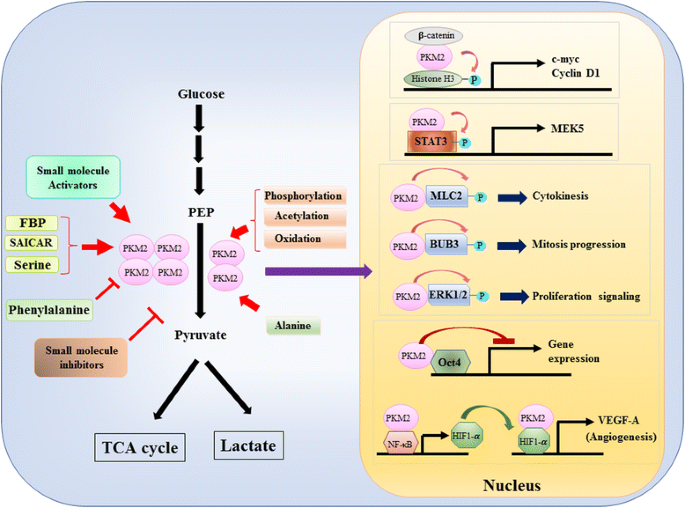
生理学的分子および翻訳後修飾によるPKM2活性の調節および転写調節におけるPKM2の潜在的な役割
第二のエピソード:シグナリングとしてPKM2細胞質におけるモジュレーター
解糖酵素としての機能に加えて、pkm2は、細胞質中の相互作用するタンパク質の同定のために、より多くの細胞プロセ 例えば、PKM2は、A−Raf、Bcr−ABL融合キナーゼ、線維芽細胞増殖因子受容体1(FGFR1)などを含むいくつかのチロシンキナーゼの相互作用タンパク質であることが示さ . これらの結合パートナーは、細胞代謝を変化させるためにPKM2の二量体/四量体変化を調節することが示されている。 しかし、PKM2は、相互的にシグナル伝達を調節するために、これらの結合パートナーの触媒動力学、基質結合および細胞質の位置に影響を与える可能性が 多くの細胞内シグナリングメディエーターは、シグナル伝達のための特定のタンパク質複合体を組み立てるために蛍光体-チロシン残基に結合するこ 今日まで、PKM2結合パートナーのリストは継続的に増加しています。 我々は、いくつかの新しいメンバーを強調し、ここで彼らの生物学的含意を議論します。 Mukheriee et al. PKM2は、mRNAの安定性と翻訳効率の制御に重要な役割を果たすRNA結合タンパク質であるHuRと結合し、神経膠腫細胞の細胞周期の進行と増殖を促進するこ 興味深いことに、3′-untranslational領域(3′-UTR)でAUリッチ要素を介してmRNAの数を結合することができる別のRNA結合タンパク質トリステトラプロリンもPKM2相互作用パートナーであることが判明し、PKM2は、乳癌の成長を調節するためにトリステトラプロリンのリン酸化と分解を誘導した。 これら二つの研究は、PKM2の潜在的な翻訳制御機能を意味する。 最近、Liang et al. 新しいPKM2パートナーとして抗アポトーシスタンパク質Bcl2を同定した。 彼らは、酸化ストレスは、ユビキチン化依存性経路を介してその分解を防止することにより、リン酸化し、Bcl2を安定化したミトコンドリアへのPKM2の転座を誘導することを実証した。 これらのデータは、PKM2が癌細胞が細胞内代謝変化または細胞外侮辱によって誘発される酸化ストレスを適応させるのに役立つことを示唆した。/p>
第三話: 核における転写調節因子としてのPKM2
遺伝子転写またはエピジェネティック修飾の調節におけるPKM2の核的役割は、PKM2がY333リン酸化β-カテニンと結合し、β-カテニン-PKM2複合体がヌクレオソームに動員され、スレオニン11でヒストンH3をリン酸化することによって示唆された。 1). このリン酸化はその後、β-カテニン標的遺伝子のアップレギュレーションにつながったヒストンH3アセチル化を増加させた。 PKM2によって直接リン酸化される別の転写因子は、転写3のシグナル変換器および活性化因子(STAT3)である。 チロシン705でSTAT3のPKM2を介したリン酸化は、マイトジェン活性化プロテインキナーゼキナーゼ5(MEK5)の発現をアップレギュレートするSTAT3活性を強化した。 転写因子の横に、PKM2は、ミオシン軽鎖2(MLC2)、BUB3および細胞外シグナル調節キナーゼ1および2(ERK1およびERK2)をリン酸化することが示されている。 興味深いことに、PKM2はまた、遺伝子発現に影響を与えるためにリン酸化非依存的な方法を介して作用する。 例えば、PKM2は、幹細胞の自己再生を制御し、Oct4媒介性転写を阻害するマスター転写因子の1つであるOct4と結合することが見出されている。 PKM2はまた、核内のNF−κ BおよびHIF−1αと相互作用し、HIF−1α標的遺伝子VEGF−Aの発現を活性化することによって腫瘍血管形成を増強することができる。 これらの研究は強く、様々な生理学的および病理学的状況におけるPKM2の核局在とプロテインキナーゼ機能を示唆したが、しかし、核PKM2を介した遺伝子発現の重要性は、PKM2ノックアウト細胞を用いた研究によって挑戦されている。 標識されたPEPおよびPKM2ヌルマウス胚性線維芽細胞を使用することにより、Hosiosら(1 9 9 9)、Hosios e t a l. PEP依存性リン酸化は細胞内で一般的な事象ではなく、反応がPKM2によって触媒されないことを示した。 これらの研究の不一致は現在未解決であり、PKM2のプロテインキナーゼ活性はさらなる確認を必要とする。
第四のエピソード:細胞外シグナリングコミュニケータとしてのPKM2
細胞外PKM2の存在は、PKM2の生物学的機能の研究のための新しい道を開 Buschow et al. PKM2は、B細胞エキソソームで検出することができ、MHCクラスII関連タンパク質として同定された最初の証拠を提供しました。 その後の二つの研究はまた、PKM2は、様々な細胞によって放出されたエキソソームに存在することを示した。 現在、ExoCartaやEVpediaのようないくつかの公開データベースは、タンパク質、脂質、異なる種の細胞外小胞の核酸を含むコンポーネントの包括的な情報を提供します。 すべてのデータは、PKM2がエキソソームのパッケージタンパク質であることを確認する。 最近の研究では、宿主細胞からレシピエント細胞に異なる成分を送達することにより、エキソソームの伝達的役割を明確に実証している。 PKM2が細胞-細胞クロストークに役割を果たす可能性があることが期待される。
新たな証拠は確かにこの仮説をサポートしています。 例えば、最近の研究は、pkm2を循環させる血液が、内皮細胞の成長、遊走およびマトリックス接着を増加させることによって、腫瘍成長および血管新生を促進し得ることを実証した。 別の調査はまた、結腸癌細胞から分泌されたPKM2は、PI3K/AktおよびWnt/β-カテニン経路を活性化することにより、細胞遊走を強化するために自己分泌刺激を介して作用する可能性があることを示した。 癌細胞に加えて、組織損傷部位の好中球は、血管新生および創傷治癒を促進するためにPKM2を放出する可能性がある。 我々の最近の研究はまた、組換えPKM2タンパク質は、表皮成長因子受容体(EGFR)のリン酸化と活性化を誘導することができることを示した。 また、我々は優先的に二量体PKM2を形成したR339E変異体PKM2は、四量体PKM2よりも有意にEGFRを活性化することがわかった。 Keller et al. succinyl-5-aminoimidazole-4-carboxamide-1-ribose-5′-phosphate(SAICAR)、PKM2のプロテインキナーゼ活性を刺激することができる細胞内代謝産物とHela細胞の処理後PKM2の潜在的な基質として154タンパク質 彼らはまた、PKM2基質としてEGFRを発見した。 彼らの結果は2つの点で私たちとは異なります。 まず、我々の研究で活性化されたシグナル伝達経路は、細胞外PKM2によって誘発され、その研究で同定された分子標的は細胞内PKM2の潜在的な基質である。 第二に、我々の研究におけるERK1/2活性の増加はEGFR活性化によって開始され、その研究におけるERK1/2活性化はSAICAR/PKM2複合体によって直接刺激される。 両方の研究で観察された一つの同様の現象は、R339E変異PKM2は、腫瘍形成における二量体と四量体PKM2の明確な役割を示唆している野生型PKM2よりも 受容体チロシンキナーゼアレイを使用することにより、我々は細胞外PKM2のみ乳癌細胞における限られた成長因子受容体を活性化することを見出した( 現在、細胞外PKM2による受容体活性化の選択性は不明のままである。 さらに、R339E変異体PKM2がEGFRの活性化においてより強力である理由も明らかではない。 これらの質問に答えるためには、より多くの実験が必要です。対処すべきもう一つの優雅な質問は、遊離PKM2および小胞包装PKM2が腫瘍形成を促進する際に同様の効果を示すかどうかである(図10B)。
2). 遊離の細胞外PKM2は原形質膜に浸透することができず、成長因子受容体のような細胞表面タンパク質を介して細胞内シグナル伝達を活性化する 逆に、小胞包装PKM2は、癌および間質細胞によってエンドサイトーシスすることができ、小胞から放出されたPKM2は、細胞内機構を介して代謝および遺伝子発現に影響を与える可能性がある。 The PKM2-null cells or mice will be useful to elucidate whether these two extracellular forms of PKM2 could act synergistically or antagonistically in tumorigenesis.

The potential tumor-promoting effect of extracellular PKM2
The fifth episode: PKM2の未回答の不一致
上記の細胞ベースのデータに加えて、PKM2の発癌性の役割は、PKM2ノックアウトマウスの生成後に挑戦されています。 Israelsen et al. PKM2特異的エクソン10を削除することにより、条件付きノックアウトマウスモデルを生成しました。 驚くべきことに、PKM2の枯渇は、マウスにおけるBrca1遺伝子の損失によって駆動される腫瘍形成を加速したが、弱毒化していません。 これらのデータは、PKM2が癌細胞の増殖に必要とされないことを示した。 興味深いことに、PKM1の発現は、乳癌におけるPKM1の腫瘍抑制の役割を示唆している非増殖腫瘍細胞でのみ検出された。 さらに、PKM2ノックアウトマウスは、代謝の不均衡のために長い潜伏後に自発的に肝細胞癌を発症する発生率が高い。 これらの結果は、PKM2がin vivoで発癌性の役割を果たすという概念に対するものである。
続きのエピソード:PKM2は癌バイオマーカーであり、薬物標的ですか?
遺伝子工学マウスモデルの結果はPKM2の腫瘍促進活性を支持しないが、PKM2の過剰発現はヒト癌において普遍的に見られ、臨床転帰が悪い(表1)。 最近の2つのメタアナリシス研究もこの結論を支持した。 呉他 消化器系の固形腫瘍を有する2812人の患者のデータを分析し、16のコホート研究から得られ、PKM2の過剰発現は、胃癌、食道扁平上皮癌、肝細胞癌、胆道癌および口腔癌の全生存率の低下に関連していることを見出した。 しかし、PKM2は膵臓癌の予後因子ではない。 この知見は、PKM2の増加が膵臓癌患者の生存不良の指標であることを示す以前の研究と一致していない。 別の調査は4796のケースを含んでいた27の個々の調査からPKM2upregulationがプールされたデータのより悪い全生存、無病生存および無再発生存と相関していることを示 しかし、がんの種類によって層別化されたPKM2は、膵臓がんの生存率が低いとは予測していません。 集合的に、PKM2は固形腫瘍のほとんどの責任を負う予後マーカーであると思われる。
逆に、診断因子としてPKM2の使用は議論の余地があります。 プロテオミクス解析は、PKM2は肺癌の検出のための潜在的な診断マーカーであることを示した。 しかし、最近の研究では、PKM2は特異性が低いために肺癌の良好な診断マーカーではないことが示唆された。 同様に、PKM2単独では結腸癌のスクリーニングのための有用なマーカーであることはまずありません。 しかし、複数のマーカーの組み合わせは、癌診断のための感度と特異性を高めることができます。
PKM2の治療の可能性は、癌治療における興味深いイベントです。 一方から、PKM2の阻害は解糖を阻害し、遺伝子転写を阻害し、細胞増殖を抑制することが期待される。 したがって、PKM2阻害剤は、抗癌薬開発のための良い候補であると思われます。 ライブラリスクリーニングを使用することにより、Vander Heiden et al. PKM2阻害剤の三つの新規クラスを同定し、最も効果的な化合物がPKM2活性を阻害し、癌細胞の死を誘発することを示した。 最近、Ning e t a l. 新規ナフトキノン誘導体が強力なPKM2阻害剤であることが判明した。 一つの有効な化合物3kは、正常細胞にはほとんど有害な影響を示さなかったが、サブマイクロモル濃度で複数の癌細胞株の増殖を抑制した。 他の側から、PKM2の活性化はまた、腫瘍増殖を阻害し得る。 低活性PKM2二量体は、癌細胞の細胞質および核内の遺伝子転写における解糖を誘発する主要なアイソフォームであるため、四量体PKM2の形成を促進することができるPKM2活性化剤は、解糖をミトコンドリア経路に切り替え、遺伝子転写を減衰させるために核の侵入を減少させる可能性がある。 両方の効果は、代謝需要と腫瘍の退行につながる成長を支援するシグナル伝達を損ないます。 二つの先駆的研究は、様々なPKM2活性化剤を同定し、in vitroでその特異性を特徴づけた。 その後の研究では、PKM2活性化剤が実際に四量体形成を促進し、in vivoで腫瘍増殖を抑制することが示された。 これらの結果は、PKM2活性化剤が有望な抗癌薬である可能性が示唆された。化学療法に対する抵抗性は、癌治療の主要な閉塞である。
がん細胞の固有の化学抵抗性を克服することは、臨床研究にとって急務です。 癌細胞の化学抵抗性におけるPKM2の役割が明らかにされており、PKM2を標的とすることは、化学抵抗性癌細胞を再感作することが示されている。 最近の研究では、CD44はPKM2と相互作用し、PKM2のTyr105リン酸化を増加させることを介してPKM2活性を抑制することが示された。 CD44アブレーションは、好気性解糖からミトコンドリア呼吸へのスイッチを誘導し、活性酸素種(ROS)生産を増加させ、大腸癌細胞におけるシスプラチン感受性 PKM2活性の阻害は、解糖を抑制し、シスプラチン抵抗性を克服することが示された。 したがって、シスプラチンおよびPKM2阻害剤の組み合わせは、化学療法のための有効な戦略であり得る(図1 0A)。 3). PKM2はまた、膵臓癌細胞におけるゲムシタビン抵抗性の調節に関与することが報告されている。 Kim et al. 従ってPKM2が否定的にp38仲介されたp53リン酸化を調整し、p53のtranscriptional活動を減らし、プロapoptotic遺伝子の表現を抑制することによってgemcitabineの抵抗を促進するこ さらに、二量体PKM2を優先的に形成するR399E-PKM2の異所性発現は、ゲムシタビンに対する膵臓癌細胞の抵抗性を高める。 これらの証拠は、PKM2が非代謝機構を介して薬剤耐性の誘導に寄与することを示唆している。 二量体PKM2は主に癌細胞の転写調節因子として作用するため、四量体PKM2の形成を促進するPKM2活性化剤は、PKM2の非代謝機能を抑制し、膵臓癌細胞のゲムシタビンに対する耐性を低下させるのに有効である可能性がある。 核PKM2はまた、結腸直腸癌および肺癌におけるEGFR阻害剤の耐性に寄与することが実証されている。 Li et al. 核PKM2を介したSTAT3のリン酸化は、ゲフィチニブと核PKM2とSTAT3の相互作用の破壊に大腸癌細胞の感度を低下させることを示した細胞内のゲフィチニ さらに最近では、別の研究では、PKM2は、核に移動し、成長因子刺激時にポリADPリボース(PAR)と相互作用することを示しました。 PKM2のPAR結合活性は、PKM2の核保持および遺伝子転写のために重要であり、細胞増殖および腫瘍増殖の促進のために重要である。 彼らは、PKM2核機能の阻害がEGFR変異癌細胞の抵抗性を克服する可能性があると結論づけた。 両方の研究は、癌における薬剤耐性を媒介する核PKM2の重要な役割を指摘し、核PKM2を標的とすることは、耐性を無効にする有望な戦略である可能性が示唆された。