kinaza Pirogronianowa m2 napędza wiele aspektów komórek nowotworowych: od metabolizmu komórkowego, regulacji transkrypcyjnej do sygnalizacji pozakomórkowej
analiza biochemiczna poprzez charakterystykę aktywności enzymatycznej, która katalizuje tworzenie mleczanów z glukozy w lizatach komórkowych, ujawniła pierwszy wewnątrzkomórkowy szlak metaboliczny, szlak glikolityczny. Począwszy od oczyszczania frakcji, które zawierały aktywność glikolityczną, wielu pionierskich badaczy przyczyniło się do identyfikacji enzymów, które biorą udział w każdym etapie szlaku. Wyniki te budują naszą nowoczesną koncepcję wymiany oddychania tlenowego i beztlenowego oraz produkcji energii w różnych warunkach fizjologicznych i patologicznych.
istnienie enzymu katalizującego produkcję ATP poprzez przeniesienie grupy fosforanowej z PEP do ADP w wątrobie zostało po raz pierwszy opisane w 1934 roku . Późniejsza izolacja enzymu, znana później jako kinaza pirogronianowa (PK), wykazała różnice w dystrybucji tkankowej i kinetyce katalitycznej sugerując, że enzym ten może mieć różne izoformy . W latach 1982-1984 sklonowano różne geny PK z drożdży, kurczaka i szczura . Badanie funkcjonalne PKM2 zostało zainicjowane przez identyfikację genu kandydującego u myszy na początku lat 80 . Później Noguchi i in. wykazano, że dwie izoformy PK (PKM1 i PKM2) są kodowane przez ten sam gen PKM poprzez alternatywne splicing . U człowieka izoformy PKM są również wytwarzane za pomocą podobnego mechanizmu splicingu poprzez włączenie eksonu 9 i 10 do mRNA PKM1 i PKM2 oddzielnie .
kilka znalezisk zwróciło uwagę badacza na potencjalną rolę PKM2 w nowotworach. Po pierwsze, PKM2 jest embrionalną izoformą, która silnie wyraża się podczas rozwoju zwierząt. Jego transkrypcja jest atenuowana w wielu tkankach dorosłych, podczas gdy jest reaktywowana w guzach . Po drugie, badanie względnej obfitości PKM1 i PKM2 w tkankach prawidłowych i nowotworowych wykazało przejście z izoformy PKM1 na IZOFORMĘ PKM2 w różnych nowotworach, takich jak rak wątrobowokomórkowy . Po trzecie, zmiana splicingu mRNA z PKM1 na PKM2 jest wzmocniona przez onkogen c-Myc, co sugeruje, że komórki rakowe aktywnie angażują się w ten przełącznik, aby spełnić swoje wymagania w proliferacji i metabolizmie . Po czwarte, modulacja aktywności PKM2 przez aktywatory lub inhibitory wpływa na wzrost guza in vivo .
- pierwszy odcinek: PKM2 jako enzym metaboliczny w cytoplazmie
- drugi odcinek: pkm2 jako modulator sygnałowy w cytoplazmie
- trzeci odcinek: PKM2 jako regulator transkrypcyjny w jądrze
- czwarty odcinek: PKM2 jako pozakomórkowy komunikator sygnałowy
- The fifth episode: bez odpowiedzi rozbieżność PKM2
- ciąg dalszy: czy PKM2 jest biomarkerem raka i celem leków?
pierwszy odcinek: PKM2 jako enzym metaboliczny w cytoplazmie
ponieważ rola PKM2 w kontroli metabolicznej glikolizy w komórkach nowotworowych została szczegółowo przeanalizowana , podsumowujemy tylko trzy kluczowe różnice między katalizą mediowaną przez PKM1 i PKM2 a metabolizmem komórkowym. Pierwszą różnicą jest interakcja podjednostek. Zarówno PKM1, jak i PKM2 są białkami tetramerycznymi utworzonymi przez cztery identyczne podjednostki. Każda podjednostka (lub monomer) zawiera cztery domeny strukturalne, w tym domenę A, B, C i N-końcową. Monomer najpierw dimeryzuje się razem, a następnie dwa dimery wchodzą w interakcję poprzez interfejs dimer-dimer zaaranżowany przez domenę C monomeru, tworząc tetramer. Ponieważ PKM1 i PKM2 zawierają różne egzony w swoich mRNA, zmienia to kodowane aminokwasy w domenie C i zmienia stabilność tetrameru. W warunkach fizjologicznych PKM1 konstytutywnie organizuje się jako tetramer, podczas gdy PKM2 może istnieć w tetramerze lub dimerze. Drugą różnicą jest regulacja allosteryczna. W zależności od wewnątrzkomórkowego stężenia małych cząsteczek i metabolitów, aktywność PKM1 i PKM2 może być różnie regulowana. Jednym z najbardziej znanych regulatorów allosterycznych jest fruktozo-1,6-bisfosforan (FBP). Ten glikolityczny związek pośredni bezpośrednio wiąże PKM2 i zwiększa powinowactwo PKM2 do PEP . Wręcz przeciwnie, FBP nie wpływa znacząco na działalność PKM1. Oprócz FBP odnotowano wpływ na aktywność PKM2 innych metabolitów, aminokwasów i małych cząsteczek (Fig. 1). Jednak stężenie wymagane do aktywacji lub hamowania jest wysokie, a efekt modulacyjny jest niewielki. Czy te małe cząsteczki odgrywają ważną rolę w kontroli aktywności PKM w warunkach fizjologicznych jest nadal niejasne. Aktywność PKM2 jest również regulowana przez modyfikację potranslacyjną, taką jak fosforylacja, acetylacja i utlenianie, które sprzyjają niskiej aktywności dimerycznego PKM2 (Fig. 1). Trzecią różnicą jest produkcja energii i pośrednie wykorzystanie. Ponieważ PKM1 konstytucyjnie istnieje jako aktywny tetramer, główną funkcją biologiczną tej izoformy jest wytwarzanie ATP w celu dostarczania energii komórkowej. Jednakże PKM2, oprócz wytwarzania ATP, może przełączyć się na mniej aktywną formę dimeryczną w celu wytworzenia kilku glikolitycznych półproduktów, które mogą być użyte jako budulec do biosyntezy aminokwasów, lipidów i nukleotydów.
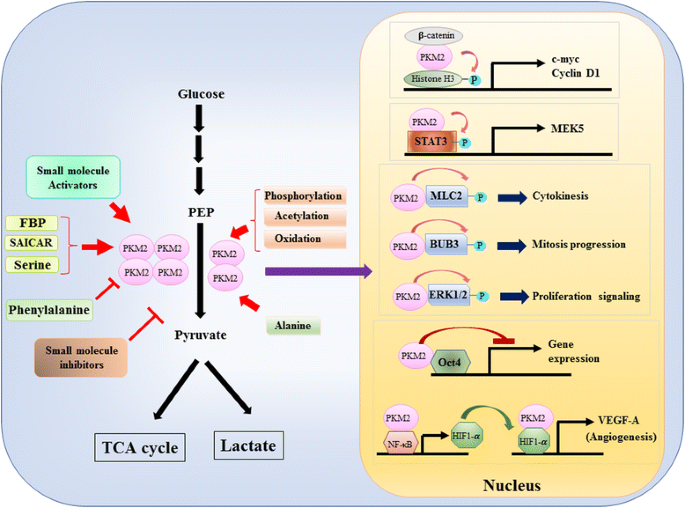
Modulacja aktywności PKM2 przez cząsteczki fizjologiczne i modyfikację potranslacyjną oraz potencjalna rola PKM2 w regulacji transkrypcji
drugi odcinek: pkm2 jako modulator sygnałowy w cytoplazmie
oprócz funkcji enzymu glikolitycznego, pkm2 proponuje się angażować w więcej procesów komórkowych ze względu na identyfikację interakcyjnych białek w cytoplazmie. Na przykład wykazano, że PKM2 jest interakcyjnym białkiem kilku kinaz tyrozynowych, w tym A-Raf, regionu klastra punktu przerwania-kinazy fuzyjnej Abelsona (BCR-ABL), receptora 1 czynnika wzrostu fibroblastów (FGFR1) itp. . Wykazano, że partnerzy wiążący modulują dimeryczną/tetrameryczną zmianę PKM2 w celu zmiany metabolizmu komórkowego. Możliwe jest jednak, że PKM2 może wzajemnie wpływać na kinetykę katalityczną, Wiązanie substratów i cytoplazmatyczną lokalizację tych partnerów wiążących w celu modulowania transdukcji sygnału. Odkrycie, że PKM2 jest białkiem wiążącym fosfor-tyrozynę wzmacnia tę możliwość, ponieważ wiele wewnątrzkomórkowych mediatorów sygnałowych może wiązać się z pozostałością fosfor-tyrozynę w celu złożenia specyficznych kompleksów białkowych do transmisji sygnału . Do tej pory lista partnerów wiążących PKM2 stale się powiększa. Wyróżniamy kilku nowych członków i omawiamy ich biologiczne konsekwencje tutaj. Mukheriee et al. wykazano, że PKM2 może wiązać się z hur, białkiem wiążącym RNA, które odgrywa ważną rolę w kontroli stabilności mRNA i wydajności translacyjnej, w celu promowania progresji cyklu komórkowego i proliferacji komórek glejaka . Co ciekawe, inne białko wiążące RNA tristetraprolina, które może wiązać szereg mRNA za pośrednictwem bogatego w AU pierwiastka w regionie 3′-nieprzetłumaczalnym (3 ’ – UTR), okazało się również partnerem oddziałującym z PKM2, a PKM2 indukuje fosforylację i degradację tristetraproliny w celu modulowania wzrostu raka piersi . Te dwa badania implikują potencjalną translacyjną funkcję kontrolną PKM2. Ostatnio liang et al. zidentyfikował anty-apoptotyczne białko Bcl2 jako nowego partnera PKM2 . Wykazali oni, że stres oksydacyjny wywołał translokację PKM2 do mitochondriów, gdzie fosforylował i stabilizował Bcl2, zapobiegając jego degradacji drogą zależną od ubikwitynacji. Dane te sugerują, że PKM2 pomaga komórkom nowotworowym w adaptacji stresu oksydacyjnego wywołanego wewnątrzkomórkową zmianą metaboliczną lub zewnątrzkomórkową zniewagą.
trzeci odcinek: PKM2 jako regulator transkrypcyjny w jądrze
jądrowa rola PKM2 w regulacji transkrypcji genów lub modyfikacji epigenetycznej została po raz pierwszy zasugerowana przez odkrycie, że PKM2 związany z β-kateniną fosforowaną Y333, a kompleks β-kateniny-PKM2 został zrekrutowany do nukleosomów do fosforylowania histonu H3 w treoninie 11 (Fig. 1). Fosforylacja ta następnie zwiększyła acetylację histonu H3, co doprowadziło do zwiększenia regulacji genów docelowych β-kateniny. Innym czynnikiem transkrypcyjnym bezpośrednio fosforylowanym przez PKM2 jest przetwornik sygnału i aktywator transkrypcji 3 (STAT3) . Mediowana przez PKM2 fosforylacja STAT3 przy tyrozynie 705 wzmocniła aktywność STAT3 w celu zwiększenia ekspresji aktywowanej mitogenem kinazy białkowej kinazy 5 (MEK5). Oprócz czynników transkrypcyjnych wykazano, że PKM2 fosforyluje miozyny łańcuch lekki 2 (MLC2), BUB3 i zewnątrzkomórkowe kinazy 1 i 2 regulowane sygnałem (ERK1 i ERK2). Co ciekawe, PKM2 działa również w sposób niezależny od fosforylacji, wpływając na ekspresję genów. Na przykład stwierdzono, że PKM2 wiąże się z Oct4, jednym z głównych czynników transkrypcyjnych, które kontrolują samoodnawianie komórek macierzystych i hamują transkrypcję za pośrednictwem Oct4 . PKM2 może również wzmocnić angiogenezę guza poprzez interakcję z NF-kB i HIF-1α w jądrze i aktywację ekspresji docelowego genu HIF-1α VEGF – A. w konsekwencji zwiększone wydzielanie VEGF – a zwiększa tworzenie naczyń krwionośnych, co przyczynia się do wzrostu guza . Chociaż badania te silnie sugerowały lokalizację jądrową i funkcję kinazy białkowej PKM2 w różnych fizjologicznych i patologicznych okolicznościach, jednak znaczenie ekspresji genów mediowanych przez pkm2 zostało zakwestionowane przez badania z wykorzystaniem komórek nokautujących PKM2. Za pomocą znakowanych PEP i PKM2-null fibroblastów zarodkowych myszy, Hosios et al. wykazano, że fosforylacja zależna od PEP nie jest częstym zdarzeniem w komórkach i reakcja nie jest katalizowana przez PKM2 . Rozbieżność tych badań jest obecnie nierozwiązana, a aktywność kinazy białkowej PKM2 wymaga dalszego potwierdzenia.
czwarty odcinek: PKM2 jako pozakomórkowy komunikator sygnałowy
obecność pozakomórkowego PKM2 otwiera nową drogę do badania funkcji biologicznej PKM2. Buschow et al. dostarczył pierwszych dowodów na to, że PKM2 może być wykryty w egzosomach komórek B i został zidentyfikowany jako białko związane z MHC klasy II . Dwa późniejsze badania wykazały również, że PKM2 istnieje w exosomach uwalnianych przez różne komórki . Obecnie kilka publicznych baz danych, takich jak ExoCarta i EVpedia, dostarcza wyczerpujących informacji na temat składników, w tym białek, lipidów, kwasów nukleinowych pęcherzyków zewnątrzkomórkowych u różnych gatunków. Wszystkie dane potwierdzają, że PKM2 jest białkiem pakietowym exosomów. Ostatnie badania wyraźnie wykazały komunikacyjną rolę exosomów poprzez dostarczanie różnych składników z komórek gospodarza do komórek biorcy . Można przypuszczać, że PKM2 może odgrywać rolę w przesłuchie komórkowym.
pojawiające się dowody rzeczywiście potwierdzają tę hipotezę. Na przykład niedawne badania wykazały, że krążąca krew PKM2 może promować wzrost guza i angiogenezę poprzez zwiększenie wzrostu, migracji i adhezji matrycy komórek śródbłonka . Inne badanie wykazało również, że PKM2 wydzielany z komórek raka jelita grubego może działać poprzez stymulację autokrynną w celu zwiększenia migracji komórek poprzez aktywację szlaków PI3K/Akt i Wnt / β-kateniny . Oprócz komórek nowotworowych neutrofile w miejscach uszkodzenia tkanek mogą uwalniać PKM2 w celu promowania angiogenezy i gojenia się ran . Nasze ostatnie badania wykazały również, że rekombinowane białko PKM2 może indukować fosforylację i aktywację receptora naskórkowego czynnika wzrostu (EGFR). Ponadto odkryliśmy, że mutant R339E pkm2, który preferencyjnie tworzy dimeryczny PKM2, aktywował EGFR znacznie bardziej niż TETRAMERYCZNY PKM2. Keller et al. zidentyfikowano 154 białka jako potencjalne substraty dla PKM2 po leczeniu komórek Hela sukcynylo-5-aminoimidazolo-4-karboksyamido-1-rybozo-5′ – fosforanem (SAICAR), metabolitem wewnątrzkomórkowym, który może stymulować aktywność kinazy białkowej PKM2 . Znaleźli również EGFR jako substrat PKM2. Ich wyniki różnią się od naszych na dwa sposoby. Po pierwsze, szlaki sygnałowe aktywowane w naszym badaniu są wywoływane przez zewnątrzkomórkowy PKM2, podczas gdy cele molekularne zidentyfikowane w ich badaniu są potencjalnymi substratami wewnątrzkomórkowego PKM2. Po drugie, zwiększenie aktywności ERK1/2 w naszym badaniu jest inicjowane przez aktywację EGFR, podczas gdy aktywacja ERK1/2 w ich badaniu jest bezpośrednio stymulowana przez kompleks SAICAR / PKM2. Jednym z podobnych zjawisk zaobserwowanych w obu badaniach jest fakt, że zmutowany PKM2 r339e aktywuje cząsteczki sygnałowe bardziej znacząco niż PKM2 typu dzikiego, co sugeruje wyraźną rolę DIMERYCZNEGO i tetramerycznego PKM2 w onkogenezie. Używając receptora kinazy tyrozynowej, odkryliśmy, że zewnątrzkomórkowy PKM2 aktywował tylko receptory ograniczonego czynnika wzrostu w komórkach raka piersi (dane nie pokazane). Obecnie selektywność aktywacji receptora przez pozakomórkowy PKM2 pozostaje nieznana. Ponadto nie jest jasne, dlaczego Mutant R339E PKM2 jest silniejszy w aktywacji EGFR. Potrzeba więcej eksperymentów, aby odpowiedzieć na te pytania.
kolejnym eleganckim pytaniem, które należy rozwiązać, jest to, czy wolny PKM2 i PKM2 opakowania pęcherzyków wykazują podobny efekt w promowaniu nowotworu (rys. 2). Wolny zewnątrzkomórkowy PKM2 nie mógł przenikać przez błonę osocza i mógł aktywować sygnalizację wewnątrzkomórkową jedynie poprzez białka powierzchniowe komórki, takie jak receptory czynnika wzrostu. I odwrotnie, PKM2 opakowania pęcherzyków może być endocytozowane przez komórki nowotworowe i zrębowe, a PKM2 uwalniane z pęcherzyków może wpływać na metabolizm i ekspresję genów poprzez mechanizm wewnątrzkomórkowy. The PKM2-null cells or mice will be useful to elucidate whether these two extracellular forms of PKM2 could act synergistically or antagonistically in tumorigenesis.

The potential tumor-promoting effect of extracellular PKM2
The fifth episode: bez odpowiedzi rozbieżność PKM2
oprócz danych opartych na komórkach omówionych powyżej, onkogenna rola PKM2 została również zakwestionowana po generacji myszy nokautujących PKM2. Israelsen et al. wygenerowano warunkowy model knockout mouse, usuwając ekson 10 specyficzny dla PKM2 . Co zaskakujące, zubożenie PKM2 przyspieszyło, ale nie atenuowało powstawania guza, spowodowane utratą genu Brca1 u myszy. Dane te wskazują, że PKM2 nie jest wymagany do proliferacji komórek nowotworowych. Co ciekawe, ekspresja PKM1 została wykryta tylko w nieroliferujących komórkach nowotworowych, co sugeruje hamującą nowotwór rolę PKM1 w raku piersi. Ponadto u myszy nokautujących PKM2 występuje duża częstość występowania raka wątrobowokomórkowego samoistnie po długim opóźnieniu spowodowanym brakiem równowagi metabolizmu . Wyniki te są sprzeczne z poglądem, że PKM2 odgrywa rolę onkogenną in vivo.
ciąg dalszy: czy PKM2 jest biomarkerem raka i celem leków?
chociaż wyniki Modelu inżynierii genetycznej myszy nie potwierdzają aktywności pkm2 promującej nowotwór, nadekspresja PKM2 jest powszechnie występująca w nowotworach ludzkich i wiąże się z słabym wynikiem klinicznym (Tabela 1) . Dwa ostatnie badania metaanalizy również potwierdziły ten wniosek. Wu et al. przeanalizowano dane 2812 pacjentów z litymi nowotworami układu pokarmowego uzyskane z 16 badań kohortowych i stwierdzono, że nadekspresja PKM2 jest związana ze zmniejszeniem całkowitego przeżycia w raku żołądka, raku płaskonabłonkowym przełyku, raku wątrobowokomórkowym, raku żółciowym i raku jamy ustnej . PKM2 nie jest jednak czynnikiem prognostycznym raka trzustki. Stwierdzenie to nie jest zgodne z wcześniejszymi badaniami wykazującymi, że wzrost PKM2 jest wskaźnikiem słabego przeżycia chorych na raka trzustki . Inne badanie obejmowało 4796 przypadków z 27 indywidualnych badań, które wykazały, że podwyższenie pkm2 jest skorelowane z gorszym przeżyciem całkowitym, przeżyciem wolnym od choroby i przeżyciem wolnym od nawrotu w zbiorczych danych . Jednak, ze względu na rodzaj nowotworu, PKM2 nie przewiduje słabego przeżycia raka trzustki. Łącznie PKM2 wydaje się być odpowiedzialnym markerem prognostycznym w większości guzów litych.
wręcz przeciwnie, zastosowanie PKM2 jako czynnika diagnostycznego jest kontrowersyjne. Analiza proteomiczna wykazała, że PKM2 jest potencjalnym markerem diagnostycznym do wykrywania raka płuc . Jednak ostatnie badania sugerują, że PKM2 nie jest dobrym markerem diagnostycznym raka płuc ze względu na niską swoistość . Podobnie samo PKM2 jest mało prawdopodobne, aby było użytecznym markerem do badań przesiewowych raka jelita grubego . Jednak połączenie wielu markerów może zwiększyć czułość i swoistość w diagnostyce raka .
potencjał terapeutyczny PKM2 jest intrygującym wydarzeniem w leczeniu nowotworów. Z jednej strony oczekuje się, że hamowanie PKM2 hamuje glikolizę, upośledza transkrypcję genów i hamuje proliferację komórkową. Dlatego inhibitory PKM2 wydają się być dobrymi kandydatami do rozwoju leków przeciwnowotworowych. Za pomocą biblioteki screening, Vander Heiden et al. zidentyfikowano trzy nowe klasy inhibitorów PKM2 i wykazano, że najskuteczniejszy związek hamuje aktywność PKM2 i indukuje śmierć komórek nowotworowych . Ostatnio Ning et al. stwierdzono, że nowe pochodne naftochinonu są silnymi inhibitorami PKM2 . Jeden skuteczny związek 3 k hamował proliferację wielu linii komórek nowotworowych w stężeniach sub-mikromolowych, podczas gdy wykazywał niewielki szkodliwy wpływ na normalne komórki. Z drugiej strony aktywacja PKM2 może również hamować wzrost guza. Ponieważ dimer PKM2 o niskiej aktywności jest główną izoformą, która wyzwala glikolizę w cytoplazmie i transkrypcję genów w jądrze w komórkach nowotworowych, aktywatory PKM2, które mogą promować tworzenie TETRAMERYCZNEGO PKM2, mogą zmienić glikolizę na szlak mitochondriów i zmniejszyć wejście jądrowe w celu osłabienia transkrypcji genów. Oba efekty upośledzają zapotrzebowanie metaboliczne i sygnalizację wspomagającą wzrost, która prowadzi do regresji nowotworu. W dwóch pionierskich badaniach zidentyfikowano różne aktywatory PKM2 i scharakteryzowano ich specyficzność in vitro . Kolejne badania wykazały, że aktywatory PKM2 rzeczywiście sprzyjały tworzeniu tetramerów i hamowały wzrost guza in vivo. Wyniki te sugerowały, że aktywatory PKM2 mogą być obiecującymi lekami przeciwnowotworowymi.
oporność na chemioterapię jest główną przeszkodą w leczeniu nowotworów. Przezwyciężenie wrodzonej chemooporności komórek nowotworowych jest pilne do badań klinicznych. Rola PKM2 w chemorezystancji komórek nowotworowych została ujawniona, a ukierunkowanie PKM2 wykazano, aby ponownie uczulać chemorezystujące komórki nowotworowe. Ostatnie badania wykazały, że CD44 wchodzi w interakcje z PKM2 i hamuje aktywność PKM2 poprzez zwiększenie fosforylacji TYR105 PKM2. Ablacja CD44 wywołała przejście od glikolizy tlenowej do oddychania mitochondrialnego i zwiększenie produkcji reaktywnych form tlenu (ROS), co spowodowało zwiększenie wrażliwości cisplatyny w komórkach raka jelita grubego . Wykazano, że hamowanie aktywności PKM2 hamuje glikolizę i pokonuje oporność na cisplatynę. Dlatego połączenie cisplatyny i inhibitorów PKM2 może być skuteczną strategią chemioterapii (rys. 3). PKM2 uczestniczy również w regulacji oporności na gemcytabinę w komórkach raka trzustki. Kim i in. wykazano, że PKM2 Promuje oporność na gemcytabinę poprzez ujemną regulację fosforylacji p53 za pośrednictwem p38, zmniejszając tym samym aktywność transkrypcyjną p53 i hamując ekspresję genów proapoptotycznych . Ponadto ektopowa ekspresja R399E-PKM2, która preferencyjnie tworzy dimeryczny PKM2, zwiększa oporność komórek raka trzustki na gemcytabinę. Te dowody sugerują, że PKM2 przyczynia się do indukcji oporności na leki poprzez mechanizm niemetaboliczny. Ponieważ dimeryczny PKM2 działa głównie jako regulator transkrypcji w komórkach nowotworowych, aktywatory PKM2, które promują tworzenie TETRAMERYCZNEGO PKM2, mogą być skuteczne w hamowaniu nie metabolicznej funkcji PKM2 i zmniejszaniu oporności na gemcytabinę w komórkach raka trzustki. Wykazano również, że jądrowy PKM2 przyczynia się do oporności inhibitora EGFR w raku jelita grubego i raka płuc . Li i in. wykazano, że fosforylacja STAT3 za pośrednictwem PKM2 zmniejsza wrażliwość komórek raka jelita grubego na gefitynib i zakłócenie interakcji nuklearnego PKM2 i STAT3 przywraca wrażliwość gefitynibu w komórkach . Niedawno inne badanie wykazało, że PKM2 translokuje się do jądra i współdziała z rybozą Poli-ADP (PAR) po stymulacji czynnika wzrostu. Aktywność wiązania par PKM2 ma kluczowe znaczenie dla retencji jądrowej i transkrypcji genów PKM2 i jest ważna dla promocji proliferacji komórek i wzrostu guza. Doszli do wniosku, że hamowanie funkcji jądrowej PKM2 może przezwyciężyć oporność zmutowanych komórek nowotworowych EGFR . Oba badania wskazywały na kluczową rolę pkm2 w mediacji oporności na leki w nowotworach i sugerowały, że ukierunkowanie pkm2 może być obiecującą strategią przezwyciężenia oporności.

PKM2 przyczynia się do proliferacji i lekooporności w komórkach nowotworowych