witamina C: środek przeciwutleniający
metabolizm redoks i właściwości przeciwutleniające witaminy C
wolne rodniki i utleniacze odgrywają podwójną rolę zarówno jako toksyczne, jak i korzystne związki, w procesach metabolicznych i w odpowiedzi na egzogenne stymulacje. Są one wytwarzane z normalnej aktywności metabolicznej lub z czynników środowiskowych (zanieczyszczenia, dym papierosowy i promieniowanie). Gdy nadmiar wolnych rodników nie może być zmiatany, ich nagromadzenie w organizmie generuje stres oksydacyjny . Stres oksydacyjny występuje, gdy tworzenie wolnych rodników przekracza zdolność ochrony przed nimi. Proces ten prowadzi do rozwoju chorób przewlekłych i zwyrodnieniowych, takich jak rak, choroby autoimmunologiczne, starzenie się, zaćma, reumatoidalne zapalenie stawów, choroby sercowo-naczyniowe i neurodegeneracyjne . Przeciwutleniacz jest cząsteczką, która zapobiega utlenianiu innych cząsteczek. Proces utleniania jest reakcją chemiczną, która wytwarza wolne rodniki, prowadząc do reakcji łańcuchowych, które mogą uszkodzić komórki. Działanie antyoksydacyjne witaminy C zostało dobrze udokumentowane . Witamina C jest silnym przeciwutleniaczem posiadającym zdolność oddawania atomu wodoru i tworząc stosunkowo stabilny wolny rodnik askorbylu. Witamina E, witamina C i β-karoten są znane jako witaminy przeciwutleniające, które sugerują zmniejszenie uszkodzeń oksydacyjnych i zmniejszenie ryzyka niektórych chorób przewlekłych. Choroby, takie jak zaburzenia sercowo-naczyniowe, są związane z nieodpowiednimi stężeniami kwasu L-askorbinowego, tokoferolu i β-karotenu w badaniach epidemiologicznych. Witamina C zwiększa również wchłanianie żelaza poprzez redukcję Fe3 + do Fe2 + ze źródeł żelaza niehemowego . W obecności jonów aktywnych redoks (żelazo, miedź) witamina C działa jako prooksydant, przyczyniając się do tworzenia rodników hydroksylowych, które mogą prowadzić do utleniania lipidów, DNA lub białek . Istnieją różne mechanizmy łagodzenia stresu oksydacyjnego i naprawy uszkodzonych makrocząsteczek. Enzymatyczne i nieenzymatyczne przeciwutleniacze odgrywają ważną rolę w usuwaniu wolnych rodników i reaktywnych form tlenu (ROS). Wykazano, że enzymy przeciwutleniające, katalaza (CAT), dysmutaza ponadtlenkowa (SOD), reduktaza glutationowa (GR), peroksydaza glutationowa (GSHpx) i, w roślinach, peroksydaza askorbinianowa (AA-px) i nieenzymatyczne przeciwutleniacze, w tym glutation (GSH) i askorbinian (ASC), podlegają znacznemu wpływowi stresu oksydacyjnego . Związki przeciwutleniające mogą zapobiegać niekontrolowanemu tworzeniu się wolnych rodników lub hamować ich reakcję z witrynami biologicznymi; również niszczenie większości wolnych rodników zależy od utleniania endogennych przeciwutleniaczy głównie poprzez usuwanie i redukcję cząsteczek . Uważa się, że witamina C jest ważnym rozpuszczalnym w wodzie przeciwutleniaczem, który neutralizuje ROS i zmniejsza stres oksydacyjny .
witamina C jest silnym środkiem redukującym i zmiatającym wolne rodniki w systemach biologicznych . Bierze udział w pierwszej linii obrony antyoksydacyjnej, chroniąc błony lipidowe i białka przed uszkodzeniem oksydacyjnym. Jako cząsteczka rozpuszczalna w wodzie witamina C może działać zarówno wewnątrz, jak i na zewnątrz komórek, i może neutralizować wolne rodniki i zapobiegać uszkodzeniom wolnych rodników. Witamina C jest doskonałym źródłem elektronów dla wolnych rodników, które szukają elektronu, aby odzyskać swoją stabilność. Witamina C może oddawać elektrony wolnym rodnikom i gasić ich reaktywność .
wykazano, że witamina C jest skutecznym środkiem usuwającym tlen i tlenki azotu, takie jak Rodnik ponadtlenkowy, nadtlenek wodoru, Rodnik hydroksylowy i tlen singletowy. Ta właściwość witaminy C ma istotne procesy w ochronie składników komórkowych przed uszkodzeniem wywołanym przez wolne rodniki. Ponadto witamina C skutecznie regeneruje antyoksydacyjną formę witaminy E poprzez redukcję rodników tokoferoxylowych. Proces ten chroni błony i inne przedziały komórki przed uszkodzeniem wywołanym przez wolne rodniki (ryc. 2). Peroksydaza askorbinianowa (APX) jest enzymem redukującym H2O2 do wody poprzez zastosowanie askorbinianu jako dawcy elektronów. Monodehydroascorbate to utleniony askorbinian, który jest regenerowany przez reduktazę monodehydroascorbate (MDAR). Rodnik monodehydroaskorbinianu szybko ulega dysproporcji do askorbinianu i dehydroaskorbinianu. Dehydroascorbate is reduced to ascorbate by dehydroascorbate reductase in the presence of GSH, yielding oxidized glutathione (GSSG). It is reduced by glutathione reductase (GR) using nicotinamide adenine dinucleotide phosphate hydrogen (NADPH) as an electron donor. Dehydroascorbate may be reduced nonenzymatically or catalyzed by proteins with dehydroascorbate reductase (DHAR) activity.
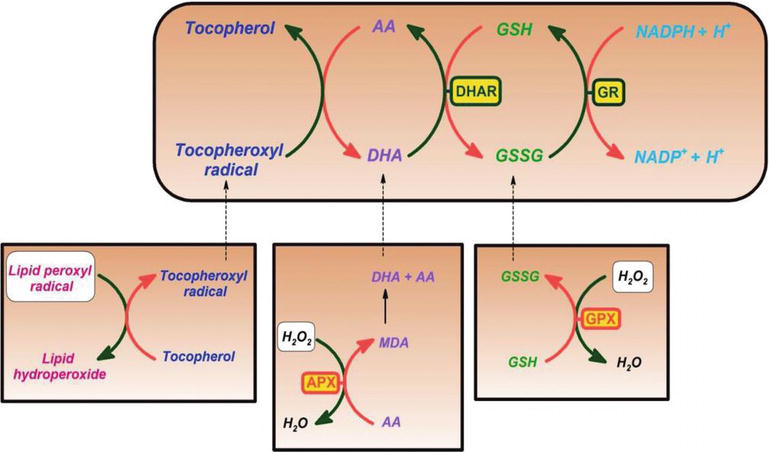
Figure 2.
xmlns:mml=”http://www.w3.org/1998/Math/MathML” xmlns:xlink=”http://www.w3.org/1999/xlink” xmlns:xsi=”http://www.w3.org/2001/XMLSchema-instanceAscorbate and redox cycling antioxidants. AA, ascorbate; DHA, dehydroascorbate; DHAR, semidehydroascorbate reductase; GSH, glutathione; GSSG, semi-glutathione reductase; GR, glutathione reductase; APX, ascorbate peroxidase; and GPX, glutathione peroxidase .
Glutathione-ascorbate cycle operates in the cytosol, mitochondria, plastids, and peroxisomes in plants . Sugeruje się, że cykl glutation-askorbinian odgrywa kluczową rolę w detoksykacji H2O2, ze względu na wysokie stężenia glutationu, askorbinianu i NADPH w komórkach roślinnych. Inne enzymy, takie jak askorbinian i peroksydazy Glutationowe, które wykorzystują tioredoksyny lub glutaredoksyny jako substraty redukujące, również odgrywają rolę w usuwaniu H2O2 w roślinach (ryc. 2).
witamina C tworzy również Rodnik półhydroaskorbylowy, stosunkowo długo żyjący Rodnik, regenerujący witaminę E z jej radykalnej postaci, a także usuwający rodniki. Komórki roślinne i zwierzęce zawierają enzym reduktazy semidehydroaskorbinianowej zależny od NADH (EC 1.6.5.4), redukujący Rodnik z powrotem do witaminy C poprzez zastosowanie NADH jako źródła środka redukującego (fig.2). Zarówno enzymatycznie, jak i nieenzymatycznie, może nieodwracalnie rozkładać się do kwasu diketoglukonowego lub może zostać przekształcony w askorbinian w reakcji zależnej od glutationu .
jako substancja redukująca i dawca elektronów, podczas usuwania wolnych rodników witamina C przekazuje wysokoenergetyczne elektrony w celu neutralizacji wolnych rodników i jest utleniona do kwasu dehydroaskorbinowego. Kwas dehydroaskorbinowy może być przekształcony z powrotem w kwas askorbinowy do ponownego użycia lub może być metabolizowany, uwalniając więcej elektronów. Chociaż witamina C jest wchłaniana z jelit przez zależny od sodu transporter witaminy C, większość komórek transportuje witaminę C w postaci utlenionej (kwas dehydroaskorbinowy) przez transporter glukozy 1. Kwas dehydroaskorbinowy jest redukowany w celu wytworzenia kwasu askorbinowego wewnątrz komórki, chroniąc mitochondria przed uszkodzeniami oksydacyjnymi wywołanymi przez wolne rodniki (fig. 2 i 3). Silnie reaktywne wolne rodniki (np., RO–, RO2–, OH–,, NO2) są redukowane przez askorbinian, a nowo wytworzony Rodnik askorbylu jest słabo reaktywny. Askorbinian może również usuwać nieradialne reaktywne gatunki, pochodzące z nadtlenoazotynów, takie jak kwas podchlorowy, ozon i środki azotujące. Witamina C jest monosacharydowym katalizatorem utleniania-redukcji (redox) występującym zarówno u zwierząt, jak i roślin. Działanie przeciwutleniające witaminy C wynika z jej zdolności do oddawania elektronów zarówno z drugiego, jak i trzeciego węgla. Podczas ewolucji naczelnych jeden z enzymów potrzebnych do wytworzenia kwasu askorbinowego został utracony przez mutację, ludzie muszą go uzyskać z diety ; większość zwierząt może syntetyzować tę witaminę w swoich ciałach i nie wymaga jej w diecie . Witamina C jest potrzebna do konwersji prokolagenu do kolagenu poprzez utlenianie pozostałości proliny do hydroksyproliny. W innych komórkach utrzymuje się w zredukowanej postaci w reakcji z glutationem . Jak pokazano na fig. 2 i 3, kwas askorbinowy jest katalizatorem redoks, który może redukować, a tym samym neutralizować ROS, takie jak nadtlenek wodoru (H2O2) (fig.2 i 3).
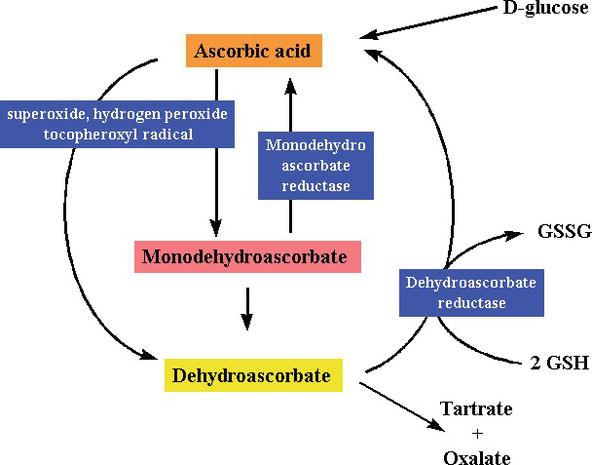
Figure 3.
xmlns:mml=”http://www.w3.org/1998/Math/MathML” xmlns:xlink=”http://www.w3.org/1999/xlink” xmlns:xsi=”http://www.w3.org/2001/XMLSchema-instanceSynthesis and degradation ofl-ascorbic acid in plant tissues .
Ascorbic acid has direct antioxidant effects, and also it is a substrate for the redox enzyme ascorbate peroxidase, that is particularly important in stress resistance in plants. Kwas askorbinowy występuje w wysokich stężeniach we wszystkich częściach roślin, zwłaszcza w chloroplastach, które osiągają tam stężenie 20 mM . Dehydroaskorbinian (DHA) i askorbinian wolnych rodników (AFR), jako półprodukt, askorbinian wolnych rodników (AFR), które są odwracalne, jednoelektronowe utlenianie powstaje z askorbinianu (Fig. Zgodnie z ogólnie przyjętym modelem enzymatycznego usuwania ROS, SOD katalizuje anion ponadtlenkowy do H2O2 i tlenu; następnie H2O2 jest redukowany do wody i tlenu molekularnego przez CAT. CAT turnover number is very high, but its affinity for H2O2 is relatively low, and consequently a certain amount of H2O2 remains in the cell.
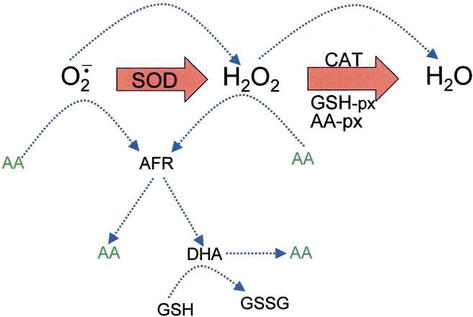
Figure 4.
xmlns:mml=”http://www.w3.org/1998/Math/MathML” xmlns:xlink=”http://www.w3.org/1999/xlink” xmlns:xsi=”http://www.w3.org/2001/XMLSchema-instanceThe role of AA in the detoxification of ROS. Blue dotted lines indicate nonenzymatic reactions.
H2O2 can react with superoxide anion formed in oxidative metabolism generating the highly reactive hydroxyl radical. GSH peroxidases (GSH-px) and AA peroxidases (AA-px) are capable of scavenging H2O2 due to their high affinity for H2O2. The cooperativity of SOD, CAT, and peroxidases ensures low amounts of superoxide anion and H2O2 and limiting the risk of hydroxyl radical formation (Figure 5).

Figure 5.
xmlns:mml=”http://www.w3.org/1998/Math/MathML” xmlns:xlink=”http://www.w3.org/1999/xlink” xmlns:xsi=”http://www.w3.org/2001/XMLSchema-instanceFoyer-Halliwell-Asada cycle .