Piruvato quinase M2 combustíveis vários aspectos das células cancerosas: a partir do metabolismo celular, transcrição regulamento de sinalização extracelular
a análise Bioquímica pela charactering a atividade enzimática, que catalisa a formação de lactato a partir da glicose na célula lisados revelou o primeiro intracelular via metabólica, o meio glicolítico. A partir da purificação de frações que continham atividade glicolítica, um número de pesquisadores pioneiros contribuiu para a identificação de enzimas que envolvem em cada passo na Via . Estes resultados constroem o nosso conceito moderno no intercâmbio de respiração aeróbica e anaeróbia e produção de energia em várias circunstâncias fisiológicas e patológicas.
a existência de uma enzima que catalisou a produção de ATP através da transferência de um grupo de fosfato de PEP para ADP no fígado foi relatada pela primeira vez em 1934 . O isolamento subsequente da enzima, conhecida como cinase de piruvato (PK) mais tarde, demonstrou diferenças na distribuição tecidular e cinética catalítica sugerindo que esta enzima pode ter isoformas diferentes . Entre 1982 e 1984, vários genes PK foram clonados a partir de levedura, frango e rato . O estudo funcional do PKM2 foi iniciado pela identificação de um gene candidato no ratinho no início da década de 1980 . Mais tarde, Noguchi et al. mostrou que duas isoformas de PK (PKM1 e PKM2) são codificadas pelo mesmo gene PKM através de splicing alternativo . Em humanos, as isoformas PKM também são produzidas através de um mecanismo de ligação semelhante, incluindo o exon 9 e 10 em mRNA PKM1 e pkm2 separadamente .vários achados chamaram a atenção do pesquisador para o potencial papel do PKM2 na tumorigénese. Em primeiro lugar, o PKM2 é a isoforma embrionária que se expressa muito durante o desenvolvimento animal. Sua transcrição é atenuada em um número de tecidos adultos, enquanto é reativado em tumores . Em segundo lugar, o estudo da abundância relativa de PKM1 e PKM2 nos tecidos normais e tumorais demonstrou uma mudança da isoforma PKM1 para a isoforma PKM2 em vários cancros como o carcinoma hepatocelular . Em terceiro lugar, a alteração do splicing do mRNA de PKM1 para PKM2 é reforçada pelo oncogeno c-Myc sugerindo que as células cancerígenas se envolvem activamente nesta mudança para se adaptarem às suas necessidades em proliferação e metabolismo . Em quarto lugar, a modulação da atividade PKM2 por ativadores ou inibidores afetam o crescimento tumoral in vivo .o primeiro episódio: PKM2 como uma enzima metabólica no citoplasma
uma vez que o papel de PKM2 no controlo metabólico da glicólise em células de câncer tem sido extensivamente analisados , nós só podemos resumir três diferenças cruciais entre PKM1 – e PKM2 mediada por catálise e metabolismo celular aqui. A primeira diferença é a interação subunitária. Tanto PKM1 quanto PKM2 são proteínas tetraméricas formadas por quatro subunidades idênticas. Cada subunidade (ou monômero) contém quatro domínios estruturais, incluindo domínios A, B, C E N-terminal. O monômero primeiro dimeriza em conjunto e, em seguida, dois dimers interagem através da interface dímero-dímero orquestrado pelo domínio C de monômero para formar um tetrâmero. Como PKM1 e PKM2 incluem diferentes exons em seus mRNAs, isso altera os aminoácidos codificados no domínio C e altera a estabilidade do tetramer. Sob condição fisiológica, PKM1 organiza-se constitucionalmente como um tetrâmero enquanto PKM2 pode existir no tetrâmero ou dímero. A segunda diferença é a regulamentação alostérica. Dependendo das concentrações intracelulares de pequenas moléculas e metabolitos, a actividade de PKM1 e PKM2 pode ser regulada diferentemente. Um dos mais conhecidos reguladores alostéricos é a frutose-1,6-bifosfato (FBP). Este intermediário glicolítico liga-se directamente ao PKM2 e aumenta a afinidade do PKM2 para a PEP . Pelo contrário, o FBP não afecta significativamente a actividade PKM1. Para além da FBP, foram relatados outros metabolitos, aminoácidos e moléculas pequenas que afectam a actividade da PKM2 (Fig. 1). No entanto, a concentração necessária para a activação ou inibição é elevada e o efeito modulatório é modesto. Ainda não é claro se estas pequenas moléculas desempenham um papel importante no controlo da actividade PKM em circunstâncias fisiológicas. A atividade PKM2 também é regulada por modificação pós-translacional, tais como fosforilação, acetilação e oxidação, que favorecem a baixa atividade do dimérico PKM2 (Fig. 1). A terceira diferença é a produção de energia e o uso intermediário. Uma vez que PKM1 existe constitucionalmente como o tetrâmero ativo, a principal função biológica desta isoforma é a geração de ATP para suprir energia celular. No entanto, PKM2, além de produzir ATP, pode mudar para a forma dimérica menos ativa para gerar vários intermediários glicolíticos que podem ser usados como blocos de construção para a biossíntese de aminoácidos, lípidos e nucleótidos.
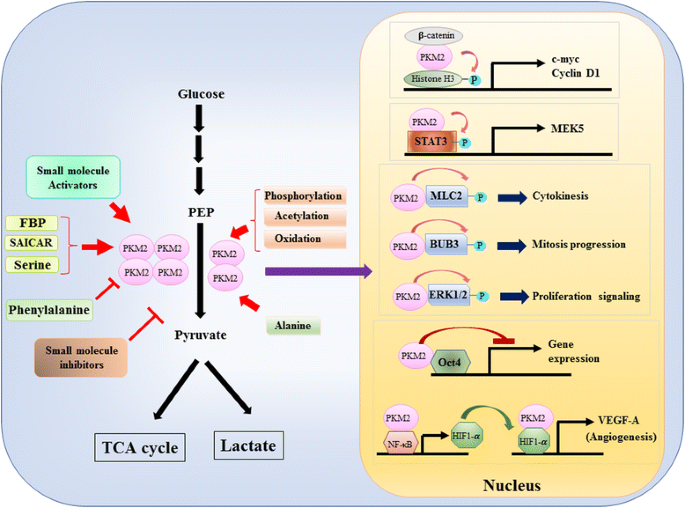
Modulação de PKM2 atividade fisiológica moléculas e pós-translacional de modificação e o papel potencial das PKM2 na regulação transcricional
segundo episódio: PKM2 como uma sinalização modulador no citoplasma
além de funcionar como um glycolytic enzima, PKM2 é proposto envolver em mais processos celulares devido à identificação de interação de proteínas no citoplasma. Por exemplo, PKM2 mostrou ser uma proteína interagindo de várias cinases tirosinas, incluindo a-Raf, Região de cluster de ponto de ruptura-Abelson (BCR-ABL) cinase de fusão, receptor 1 do fator de crescimento de fibroblast (FGFR1), etc. . Estes parceiros de ligação demonstraram modular a alteração dimérica / tetramérica de PKM2 para alterar o metabolismo celular. No entanto, é possível que o PKM2 possa afetar reciprocamente a cinética catalítica, a ligação do substrato e a localização citoplasmática destes parceiros de ligação para modular a transdução do sinal. A descoberta de que o PKM2 é uma proteína de ligação fosforetiro-tirosina reforça esta possibilidade porque muitos mediadores de sinalização intracelular podem ligar-se ao resíduo fosforetirosina para montar complexos proteicos específicos para a transmissão do sinal . Até à data, a lista de parceiros de ligação PKM2 cresce continuamente. Destacamos vários novos membros e discutimos suas implicações biológicas aqui. Mukheriee et al. demonstrou que o PKM2 pode ligar-se ao HuR, Uma proteína de ligação ao ARN que desempenha um papel importante no controlo da estabilidade do mRNA e da eficiência translacional, para promover a progressão do ciclo celular e a proliferação de células do glioma . Curiosamente, descobriu-se que outra proteína de ligação ao ARN, a tristetraprolina, que poderia ligar um número de ARN através do elemento rico em UA na Região 3′-não-transferacional (3′ – UTR), também era um parceiro de interação com o PKM2, e que o PKM2 induziu fosforilação e degradação do tristetraprolin para modular o crescimento do câncer de mama . Estes dois estudos implicam uma função de controlo translacional potencial da PKM2. Recentemente, Liang et al. identificou a proteína anti-apoptótica Bcl2 como um novo parceiro PKM2 . Eles demonstraram que o stress oxidativo induziu a translocação de PKM2 para mitocôndria, onde fosforilado e estabilizado Bcl2, impedindo a sua degradação através da via dependente da ubiquitinação. Estes dados sugerem que o PKM2 ajuda as células cancerígenas a adaptar o stress oxidativo provocado pela alteração metabólica intracelular ou pelo insulto extracelular.o terceiro episódio: PKM2 como um regulador transcricional no núcleo
Um papel nuclear de PKM2 na regulação do gene de transcrição ou de modificação epigenética, foi sugerida pela constatação de que PKM2 vinculado com Y333-phosphorlated β-catenin, e o β-catenin-PKM2 complexo foi recrutado para o nucleosomes para phosphorylate da histona H3 na treonina 11 (Fig. 1). Esta fosforilação aumentou subsequentemente a acetilação H3 da histona, o que levou à regulação dos genes-alvo da β-catenina. Outro fator de transcrição diretamente fosforilado por PKM2 é o transdutor de sinal e ativador da transcrição 3 (STAT3) . Fosforilação do STAT3 mediada pela PKM2 na tirosina 705 potenciou a actividade do STAT3 para elevar a expressão da proteína activada pelo mitogénio cinase 5 (MEK5). Para além dos factores de transcrição, demonstrou-se que o PKM2 fosforila a cadeia de luz de miosina 2 (MLC2), a BUB3 e a cinase com regulação do sinal extracelular 1 e 2 (ERK1 e ERK2) . Curiosamente, PKM2 também atua via fosforilação-forma independente para afetar a expressão do gene. Por exemplo, pkm2 foi encontrado para se ligar com Oct4, um dos fatores de transcrição mestre que controlam a auto-renovação das células-tronco, e inibem a transcrição mediada por Oct4 . PKM2 também pode melhorar a angiogênese tumoral, interagindo com NF-kB e HIF-1α no núcleo e ativando a expressão do gene alvo HIF-1α VEGF-A. consequentemente, o aumento da secreção de VEGF-A aumenta a formação de vasos sanguíneos que contribui para o crescimento do tumor . Embora estes estudos sugiram fortemente a localização nuclear e a função da proteína cinase do PKM2 em várias circunstâncias fisiológicas e patológicas, no entanto a importância da expressão genética mediada pelo pkm2 nuclear tem sido contestada por estudos usando células nocaute do PKM2. Usando fibroblastos embrionários, Hosios et al. mostrou que PEP dependente de fosforilação não é um evento comum em células e a reação não catalisada por PKM2 . A discrepância destes estudos está actualmente por resolver e a actividade da proteína cinase do PKM2 necessita de confirmação adicional.
o quarto episódio: PKM2 como um comunicador de sinalização extracelular
a presença de PKM2 extracelular abre uma nova via para o estudo da função biológica PKM2. Buschow et al. forneceu a primeira prova de que a PKM2 podia ser detectada em exossomas de células B e foi identificada como uma proteína associada à MHC classe II. Dois estudos subsequentes também indicaram que o PKM2 existe em exossomas liberados por várias células . Atualmente, vários bancos de dados públicos como ExoCarta e EVpedia fornecem informações completas para os componentes, incluindo proteínas, lípidos, ácidos nucleicos de vesículas extracelulares em diferentes espécies. Todos os dados confirmam que PKM2 é uma proteína de embalagem de exossomas. Estudos recentes demonstraram claramente um papel comunicativo dos exossomas através da entrega de diferentes componentes das células hospedeiras às células receptoras . É previsível que o PKM2 possa desempenhar um papel no crosstalk das células.a evidência emergente de fato suporta esta hipótese. Por exemplo, um estudo recente demonstrou que o sangue circulante PKM2 pode promover o crescimento tumoral e angiogênese, aumentando o crescimento, migração e adesão matriz das células endoteliais . Outra investigação também mostrou que o PKM2 secretado das células cancerígenas do cólon pode actuar através de uma estimulação autocrina para aumentar a migração celular activando as vias PI3K/Akt e Wnt/β-catenina . Além das células cancerígenas, os neutrófilos nos locais de lesão dos tecidos podem libertar PKM2 para promover a angiogénese e a cicatrização das feridas . O nosso estudo recente também demonstrou que a proteína pkm2 recombinante pode induzir fosforilação e activação do receptor do factor de crescimento epidérmico (EGFR) . Além disso, descobrimos que o r339e mutante PKM2 que preferencialmente formou o dimeric PKM2 ativado EGFR mais significativamente do que o tetramérico PKM2. Keller et al. identificou 154 proteínas como potenciais substratos do PKM2 após tratamento de células Hela com succinil-5-aminoimidazol-4-carboxamida-1-ribose-5′-fosfato (SAICAR), um metabolito intracelular que pode estimular a actividade da proteína cinase do PKM2 . Eles também encontraram o EGFR como um substrato PKM2. Os resultados deles são diferentes dos nossos de duas maneiras. Em primeiro lugar, as vias sinalizadoras activadas no nosso estudo são elicitadas pelo PKM2 extracelular, enquanto os alvos moleculares identificados no seu estudo são substratos potenciais do pkm2 intracelular. Em segundo lugar, o aumento da actividade ERK1/2 no nosso estudo é iniciado por activação EGFR enquanto a activação ERK1/2 no seu estudo é directamente estimulada pelo complexo SAICAR/PKM2. Um fenômeno similar observado em ambos os estudos é que o r339e mutante PKM2 ativa moléculas sinalizadoras mais significativamente do que o tipo selvagem PKM2 sugerindo o papel distinto do dimérico e tetramérico PKM2 na oncogênese. Usando o receptor da matriz de tirosina cinase, descobrimos que o PKM2 extracelular apenas ativou os receptores limitados do fator de crescimento nas células do câncer de mama (dados não mostrados). Atualmente, a seletividade da ativação do receptor por PKM2 extracelular permanece desconhecida. Além disso, também não é claro por que razão o pkm2 mutante R339E é mais potente na activação do EGFR. São necessárias mais experiências para responder a estas perguntas.outra questão elegante a ser abordada é a de saber se o PKM2 livre e o pkm2 em embalagens de vesículas apresentam um efeito semelhante na promoção da tumorigénese (Fig. 2). O PKM2 extracelular livre não conseguia penetrar na membrana plasmática e só podia activar a sinalização intracelular através das proteínas da superfície celular, como os receptores do factor de crescimento. Inversamente, a embalagem vesicular PKM2 pode ser endocitada por células cancerígenas e estroma, e a PKM2 libertada das vesículas pode afectar o metabolismo e a expressão genética através do mecanismo intracelular. The PKM2-null cells or mice will be useful to elucidate whether these two extracellular forms of PKM2 could act synergistically or antagonistically in tumorigenesis.

The potential tumor-promoting effect of extracellular PKM2
The fifth episode: discrepância sem resposta de PKM2
além dos dados baseados em células discutidos acima, o papel oncogénico de PKM2 também foi questionado após a geração de ratos nocaute PKM2. Israelsen et al. gerou um modelo condicional de knockout do mouse ao apagar o exon 10 específico do PKM2 . Surpreendentemente, a depleção do PKM2 acelerou, mas não atenuou, a formação tumoral provocada pela perda do gene Brca1 em ratinhos. Estes dados indicaram que o PKM2 não é necessário para a proliferação de células cancerígenas. Curiosamente, a expressão PKM1 só foi detectada em células tumorais não proliferantes sugerindo um papel supressivo do pkm1 no câncer de mama. Além disso, os ratinhos Nocaute da PKM2 têm uma elevada incidência de desenvolvimento espontâneo de carcinoma hepatocelular após uma latência prolongada devido ao desequilíbrio do metabolismo . Estes resultados contra a noção de que o PKM2 desempenha um papel oncogênico in vivo.o episódio continua: o PKM2 é um biomarcador cancerígeno e alvo de drogas?
embora os resultados do modelo do rato geneticamente modificado não apoiem a actividade de promoção tumoral da PKM2, a sobre-expressão da PKM2 é universalmente encontrada em cancros humanos e está associada a resultados clínicos deficientes (Tabela 1) . Dois estudos recentes de meta-análise também apoiaram esta conclusão. Wu et al. analisou os dados de 2812 doentes com tumores sólidos do aparelho digestivo obtidos a partir de 16 estudos de coorte e concluiu que a sobreexpressão do PKM2 está associada à redução da sobrevivência global no cancro gástrico, carcinoma esofágico de células escamosas, carcinoma hepatocelular, cancro biliar e cancro oral . No entanto, o PKM2 não é um factor de prognóstico para o cancro pancreático. Este achado não é consistente com estudos anteriores mostrando que o aumento da PKM2 é um indicador da baixa sobrevivência dos doentes com cancro pancreático . Uma outra investigação incluiu 4796 casos de 27 estudos individuais, que demonstraram que o upregulation do PKM2 está correlacionado com o agravamento da sobrevivência global, a sobrevivência livre de doença e a sobrevivência livre de recorrência nos dados agrupados . No entanto, estratificado por tipo de cancro, O PKM2 não prevê uma fraca sobrevivência do cancro pancreático. Coletivamente, PKM2 parece ser um marcador prognóstico responsável na maioria dos tumores sólidos.
Em contrário, o uso de PKM2 como um fator de diagnóstico é controverso. Uma análise proteômica demonstrou que PKM2 é um potencial marcador de diagnóstico para a detecção de câncer de pulmão . No entanto, um estudo recente sugeriu que o PKM2 não é um bom marcador de diagnóstico para o câncer de pulmão devido à baixa especificidade . Do mesmo modo, é pouco provável que o PKM2, por si só, seja um marcador útil para o rastreio do cancro do cólon . No entanto, a combinação de Marcadores Múltiplos pode aumentar a sensibilidade e especificidade para o diagnóstico de cancro .o potencial terapêutico da PKM2 é um acontecimento intrigante no tratamento do cancro. De um lado, espera-se que a inibição da PKM2 iniba a glicólise, compromete a transcrição genética e suprime a proliferação celular. Portanto, os inibidores PKM2 parecem ser bons candidatos para o desenvolvimento de medicamentos anti-câncer. Usando a exibição da biblioteca, Vander Heiden et al. identificou três novas classes de inibidores da PKM2 e mostrou que o composto mais eficaz inibiu a actividade da PKM2 e induziu a morte de células cancerígenas . Recentemente, Ning et al. descobri que os novos derivados da naftoquinona são potentes inibidores da PKM2 . Um composto eficaz 3 k suprimiu a proliferação de várias linhas celulares cancerígenas em concentrações sub-micromolares, enquanto mostrou pouco efeito prejudicial sobre as células normais. Do outro lado, a ativação do PKM2 também pode inibir o crescimento do tumor. Devido a baixa atividade PKM2 dímero é a principal isoforma que aciona a glicólise no citoplasma e do gene de transcrição no núcleo de células de câncer, PKM2 ativadores, que pode promover a formação de tetrameric PKM2 pode mudar a glicólise para a mitocôndria via e reduzir nuclear entrada para atenuar a gene transcrição. Ambos os efeitos prejudicam a demanda metabólica e a sinalização de suporte ao crescimento que leva à regressão do tumor. Dois estudos pioneiros identificaram vários ativadores PKM2 e caracterizaram sua especificidade in vitro . Um estudo subsequente demonstrou que os ativadores PKM2 realmente promoveram a formação de tetrâmeros e suprimiram o crescimento tumoral in vivo . Estes resultados sugerem que os ativadores PKM2 podem ser promissores contra o câncer.a resistência à quimioterapia é um grande bloqueio no tratamento do cancro. Superar a quimiorresistência inerente das células cancerosas é urgente para a pesquisa clínica. Os papéis do PKM2 na quimiorresistência de células cancerosas foram revelados, e o alvo do PKM2 tem sido mostrado para reensibilizar células cancerosas resistentes à quimioterapia. Um estudo recente mostrou que o CD44 interage com o PKM2 e suprime a actividade do PKM2 através do aumento da fosforilação Tir105 do PKM2. A ablação CD44 induziu a mudança da glicólise aeróbica para a respiração mitocondrial e o aumento da produção de espécies reativas de oxigênio (ROS), resultando no aumento da sensibilidade da Cisplatina nas células do câncer colorectal . A inibição da actividade da PKM2 demonstrou suprimir a glicólise e superar a resistência à cisplatina. Assim, a associação de cisplatina e inibidores da PKM2 pode ser uma estratégia eficaz para a quimioterapia (Fig. 3). Foi também referido que o PKM2 participa na regulação da resistência à gemcitabina nas células cancerígenas pancreáticas. Kim et al. demonstrou que a PKM2 promove a resistência à gemcitabina através da regulação negativa da fosforilação p53 mediada pela p38, reduzindo assim a actividade transcritional da p53 e suprimindo a expressão de genes pro-apoptóticos . Além disso, a expressão ectópica do R399E-PKM2, que forma preferencialmente o dimérico PKM2, aumenta a resistência das células cancerígenas pancreáticas à gemcitabina. Estas evidências sugerem que o PKM2 contribui para a indução da resistência ao fármaco através de um mecanismo não metabólico. Uma vez que o dimeric PKM2 actua principalmente como um regulador transcritional nas células cancerígenas, os activadores do PKM2 que promovem a formação do pkm2 tetramérico podem ser eficazes na supressão da função não metabólica do PKM2 e na redução da resistência à gemcitabina nas células cancerígenas pancreáticas. Demonstrou-se também que o PKM2 Nuclear contribui para a resistência do inibidor do EGFR no cancro colorectal e no cancro do pulmão . Li et al. demonstrou que a fosforilação STAT3 mediada pelo PKM2 nuclear reduz a sensibilidade das células cancerígenas colorectais ao gefitinib e a perturbação da interacção da sensibilidade do pkm2 nuclear e do STAT3 restabeleceu a sensibilidade do gefitinib nas células . Mais recentemente, outro estudo demonstrou que o PKM2 translocata para o núcleo e interage com a Poli-ADP ribose (PAR) com a estimulação do factor de crescimento. A atividade de ligação PAR do PKM2 é fundamental para a retenção nuclear e transcrição genética do PKM2 e é importante para a promoção da proliferação celular e crescimento tumoral. Concluíram que a inibição da função nuclear PKM2 pode ultrapassar a resistência das células cancerígenas mutadas pelo EGFR . Ambos os estudos apontaram um papel crucial do pkm2 nuclear na mediação da Resistência a drogas em cancros e sugeriu que o alvo do pkm2 nuclear pode ser uma estratégia promissora para superar a resistência.

PKM2 contribui para a proliferação e resistência às drogas em células de câncer