Gliaceller I Retina Av Helga Kolb
Helga Kolb
Tre grunnleggende typer gliaceller finnes I den menneskelige retina, Muller-celler, astroglia og microglia. Alle ble beskrevet for netthinnen Av Cajal for mer enn hundre år siden (1892).
1. Muller celler.
Muller-celler er den viktigste glialcellen i netthinnen. De danner arkitektoniske støttestrukturer som strekker seg radialt over tykkelsen av netthinnen og er grensene for netthinnen på henholdsvis ytre og indre begrensende membran. En fullstendig forståelse av formen På En Muller celle er best sett Etter Golgi farging som vist opprinnelig Av Cajal (1892) nedenfor.
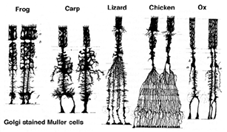
Fig. 1. Golgi-fargede Mullerceller sitter i det indre atomlaget og projiserer uregelmessig tykke og tynne prosesser i begge retninger til den ytre begrensningsmembranen og til den indre begrensningsmembranen. Muller celleprosesser insinuerer seg mellom cellelegemer av nevronene i de nukleare lagene og konvoluttgrupper av nevrale prosesser i plexiformlagene (Fig. 1). Faktisk er retinale nevrale prosesser bare tillatt direkte kontakt, uten innhyllende Mullercelleprosesser, ved deres synapser.en enkelt stamcelle gir opphav til Både Muller-celler og retinale nevroner(Turner og Cepko, 1987), men tilsynelatende i to faser. De tidligste faseneuronene født ved apikalmarginen av nevroepitelet ved siden av pigmentepitelet produserer primære nevroner som består av kegleceller, horisontale celler og ganglionceller (Fig. 2, høyre). Den andre fasen av celler som også er født ved apikale marginer, produserer Muller-celler og stav-fotoreceptorer, bipolare celler og amakrinceller (Reichenbach og Robinson, 1995) (Fig. 2, venstre). Alle de utviklende nevronene og Muller-cellene må migrere innover til sin endelige posisjon, og Det antas At Muller-celleprosessene og trunksene styrer mye av nevronmigrasjonene og styrer neurittforskjellene.
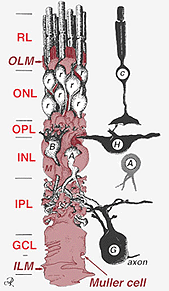
Fig. 2. 3-D skjematisk tegning av forholdet Mellom Muller celle og andre retinal nevroner
kryssene som danner den ytre begrensende membranen er Mellom Muller celler og Andre Muller celler og fotoreseptorceller som robuste desmosomer eller zonula adherens. I noen arter gap veikryss (spesialiserte membran foreninger og kanaler som tillater passasje av små molekyler og ioner) eller trange veikryss er en del av Disse Muller celle veikryss (Miller og Dowling, 1970), men ikke så i pattedyr arter der ingen fargestoff kobling har noen gang blitt observert (Robinson et al.(1993; Reichenbach Og Robinson, 1995). Overflaten Av Muller-cellen som vender mot pigmentepitelet og subretinalrommet utvides av mange fremspring Av Mullercellemembranen kjent som apikale villi. Den indre begrensende membranen, derimot, dannes av Den koniske endefoten Til Muller-cellen, men ingen spesialiserte veikryss er sett her. Muller celler danner også endfeet på de store retinale blodkarene på den indre overflaten av netthinnen. Overflaten Av Mullercellemembranen som vender mot glassplaten, er dekket med et mukopolysakkaridmateriale og danner dermed en ekte kjellermembran.Muller-celler inneholder glykogen, mitokondrier og mellomliggende filamenter som er immunoreative for vimentin og til en viss grad til glialfibrillært syreprotein (GFAP). Sistnevnte filamenter er normalt i den indre halvdelen av retinal Muller-cellene og deres endfeet (Fig. 3, venstre), men etter traumer til netthinnen som retinal detachment, blir både vimentin og GFAP massivt oppregulert og funnet gjennom cellen (Fig . 3, høyre) (Guerin et al.(1990; Fisher Og Lewis, 1995).
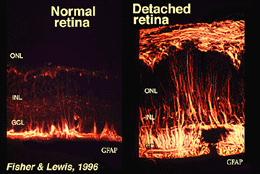
Fig. 3. GFAP immunoreaktivitet I Muller celler
Muller celler har en rekke funksjoner som alle er avgjørende for helsen til retinale nevroner. Muller-celler fungerer i et symbiotisk forhold til nevronene (For en utmerket anmeldelse se Reichenbach og Robinson, 1995). Dermed Muller cellefunksjoner inkluderer:
- Leverer endprodukter av anaerob metabolisme (nedbrytning av glykogen) til drivstoff aerob metabolisme i nervecellene.
- de mopp opp nevrale avfallsprodukter som karbondioksid og ammoniakk og resirkulere brukte aminosyre sendere.De beskytter nevroner mot eksponering for overflødige nevrotransmittere som glutamat ved hjelp av velutviklede opptaksmekanismer for å resirkulere denne senderen. De er spesielt preget av tilstedeværelsen av høye konsentrasjoner av glutaminsyntase.
- De kan være involvert i både fagocytose av nevronalt rusk og frigjøring av nevroaktive stoffer som GABA, taurin og dopamin.de antas å syntetisere retinsyre fra retinol (retinsyre er kjent for å være viktig i utviklingen av øyet og nervesystemet) (Edwards, 1994) De kontrollerer homeostase og beskytter nevroner mot skadelige endringer i deres ioniske miljø ved å ta opp ekstracellulær K+ og omfordele den.de bidrar til genereringen av electroretinogram (erg) b-bølgen (Miller Og Dowling, 1970; Newman Og Odette, 1984), den langsomme p3-komponenten AV erg (Karwoski Og Proenza, 1977) og scotopic threshold response (Str) (Frishman Og Steinberg, 1989). De gjør det ved regulering Av k + distribusjon over retinal glasslegemet grensen, over hele netthinnen og lokalt i det indre plexiform lag av netthinnen (Fig. 4, Fra Reichenbach Og Robinson, 1995, tilpasset Fra Newman, 1989).

Fig. 4. Regulering Av K + Av Muller celler i Muller celler
2. Astrocytter.Astrocytter er ikke glialceller i retinal neuroepithelium, men går inn i utviklende retina fra hjernen langs utviklende optisk nerve (Stone And Dreher, 1987; Chan-Ling 1994). De har en karakteristisk morfologi av en flatt cellekropp og en fibrøs serie utstrålende prosesser. Mellomliggende filamenter fyller sine prosesser og dermed flekker de dramatisk med antistoffer mot GFAP (Schnitzer, 1988). Astrocytcellekropper og prosesser er nesten helt begrenset til nervefiberlaget i netthinnen. Deres morfologi endres fra periferien til optisk nervehodet: fra en symmetrisk stellatform i perifer retina (Fig. 5a og b) (Schitzer, 1988) til ekstremt langstrakt nær optisk nerve (Fig. 6 og 7).
 Fig. 5a. Astrocytter i perifer retina. Schnitzer, 1988 |
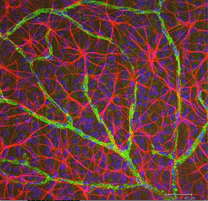 Fig. 5b. Astrocytter (rød) i rottehinnen blant ganglionceller (blå) og assosiert med blodkar (grønn). Hilsen Av Laura Fernandez-Sanchez Og Nicolas Cuenca |
i immunocytokjemisk farging (Fig. 5b) OG i HRP intracellulære injeksjoner (Fig. 7) farget astrocytter tydelig viser prosesser justert langs ganglion celle axons løpe gjennom nerve fiber laget. I distribusjon når astrocytter sin topp på optisk nervehodet og har en ganske jevn nedgang i tetthet i utstrålende ringer fra nervehodet. De er ikke til stede i avaskulær fovea eller ora serrata.
 Fig. 6. Astrocytter i sentral retina. Schnitzer, 1988 |
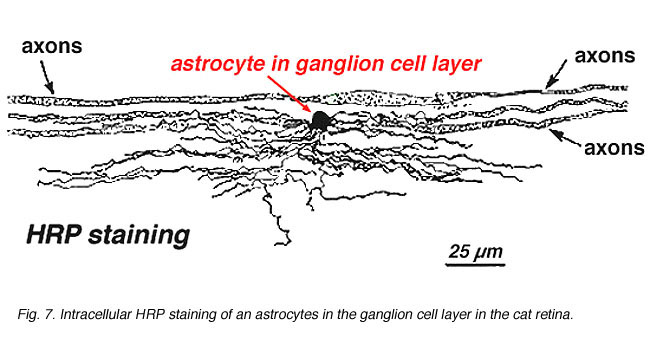 Fig. 7. Astrocytter i cat retina farget av intracellulær injeksjon AV HRP. Hilsen Av Ralph Nelson. |
Tykke og tynne astrocytter har blitt skilt ut på grunnlag av farging med antistoffer MOT GFAP (Trevino et al., 1996). Dermed astrocytter er anordnet over overflaten av ganglion celle axon bunter som de kurs i synsnerven hodet danner et rør gjennom hvilket axons kjøre (Fig. 8). Gapkryss og zonula adherenskryss har blitt beskrevet mellom astrocytiske prosesser i cat retina (H@llander et al., 1991).
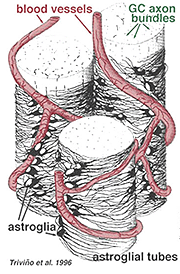
Fig. 8. 3-D-blokk av astrocytter arrangert over overflaten av ganglioncelleaksonbunter
blodkarene som går inn og blant ganglioncellebuntene, dekkes også av begge prosesser og til og med en sporadisk cellekropp av en astrocyt. Funksjonen til astrocytter som omslutter ganglioncelleaksoner og forholdet til blodkar i nervefiberlaget antyder at de er aksonale og vaskulære glialskjeder og en del av en blod-hjernebarriere. I likhet Med Muller-celler, er de kjent for å inneholde rikelig glykogen, og de kan danne en næringstjeneste for å gi glukose til nevronene. I tillegg tjener de sannsynligvis en rolle i ionisk homeostase ved regulering av ekstracellulære kaliumnivåer og metabolisme av nevrotransmittere som GABA.
3. Mikrogliale celler.den tredje glialcelletypen er angivelig av mesodermal opprinnelse og er derfor strengt tatt ikke neurogliale som astrocytene og Muller-cellene. De går inn i netthinnen sammenfallende med mesenkymale forløpere av retinale blodkar i utvikling (Chan-Ling, 1994). Mikrogliale celler er allestedsnærværende i den menneskelige retina som finnes i hvert lag av netthinnen.
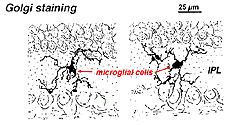
I Golgi-farget netthinnen ser de ut som merkelige, multipolare former med småcellekropper og uregelmessige korte prosesser. Faktisk, I Golgi-preparater har De noen Ganger blitt forvekslet med nerveceller, spesielt når de ligger i et nukleært lag med en enkelt orientering av deres prosesser i plexiformlaget.
Fig. 9. Golgi-farging av mikroglialceller
Mikroglialceller kan være av to typer. En form antas å komme inn i netthinnen i tidlige stadier av utvikling fra optisk nerve mesenkym og ligge sovende i retinale lag for mye av livet til netthinnen. Den andre formen for microglia ser ut til å være blodbårne celler, mulig opprinnelse fra fartøy pericytes (Boikott og Hopkins, 1981; Gallego, 1986). Begge typer kan stimuleres til en makrofagisk funksjon etter traumer til netthinnen, og deretter engasjerer de seg i fagocytose av degenererende retinale nevroner.

Fig. 10a. Lektin-farget mikroglialcelle Fra Chan-Lin, 1994.
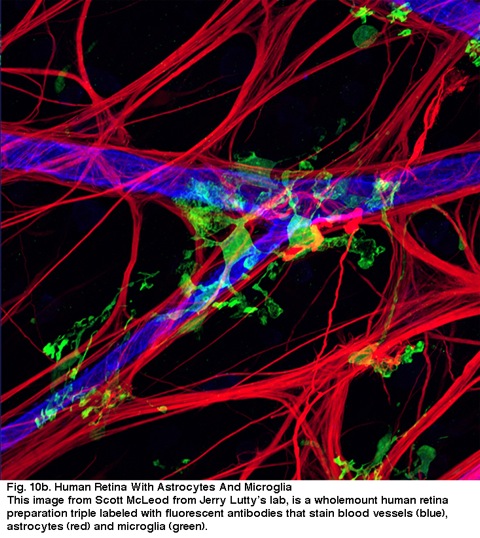
Fig. 10b. human retina med Astrocytter og Microglia. Dette bildet fra Scott McLeod Fra Jerry Lutty ‘ s lab, er en fullmount human retina forberedelse trippel laneled med fluorescerende antistoffer som flekker blodkar (blå), astrocytter (rød) og microglia (grønn).
4. Referanse.Boikott BB, Hopkins JM. Mikroglia i netthinnen av ape og andre pattedyr; dens skille fra andre typer glia og horisontale celler. Nevrovitenskap.1981;6:679–688.
Cajal SR. In: Thorpe SA, Glickstein M, translators. 1892. Strukturen av netthinnen. Springfield (IL): Thomas; 1972.
Chan-Ling T. Glial, nevronale og vaskulære interaksjoner i pattedyrets retina. Prog. Ret. Øye Res. 1994; 13: 357-389.
Edwards RB. Biosyntese av retinsyre Ved Mü glialceller: en modell for sentralnervesystemet? Prog. Ret. Øye Res. 1994; 13: 231-242.
Fisher SK, Lewis GP. Fotoreceptorer og utover: cellulære og molekylære effekter av retinal detachment.2nd Great Basin Visuell Vitenskap Symposium, II, University Of Utah Press. 1995
Frishman LJ, Steinberg RH. Lys fremkalt økning i o i proksimal del av den mørketilpassede katthinnen. J Nevrofysiol. 1989;61:1233–1243.
Gallego A. Sammenlignende studier på horisontale celler og et notat på mikroglialceller. Prog. Ret. Res. 1986; 5: 165-206.
Guerin CJ, Anderson DH, Fisher SK. Endringer i intermediær filamentimmunmerking forekommer som respons på retinal detachment og reattachment i primater.Investere. Ophthal. Vis. Sci. 1990;31:1474–1482. Karwoski CJ, Proenza LM. Forholdet Mellom Muller celle responser, en lokal transretinal potensial, og kalium flux. J Nevrofysiol. 1977;40:244–259.
Miller RF, Dowling JE. Intracellulære responser Av Muller (glial) celler i mudpuppy retina: deres forhold til b-bølgen av elektroretinogrammet. J Nevrofysiol.1970;33:323–341.
Newman EA, Odette LL. Modell av electroretinogram b-bølge generasjon: en test Av k + hypotesen. J Nevrofysiol. 1984;51:164–182.
Newman EA. Elektrofysiologi av retinale glialceller. Prog. Ret. Res. 1989; 8: 153-172.
Reichenbach A, Robinson SR. Involvering Av Mü-celler i ytre retina. In: Djamgoz MBA, Archer SN, Vallerga S, editors. Nevrobiologi og kliniske aspekter av ytre retina. London :Chapman & Hall; 1995. s. 395-416.Robinson SR, Hampson ECGM, Munro MN, Vaney DI. Enveis kobling av gapskryss mellom nevrologi. Proc. Austr. Neurosci. Soc. 1993;3:167.
Schnitzer J. Astrocytter i pattedyr retina. Prog. Ret. Res. 1988; 7: 209-232.
Stein J, Makarov F, Hollander H. glial ensheathment av soma og axon hillock av retinal ganglion celler. Vis Neurosci. 12:273–279.
Stein J, Dreher Z. Forholdet mellom astrocytter, ganglionceller og vaskulatur av netthinnen. J Comp Neurol. 1987;255:35–49. Trivino A, Ramirez JM, Salazar JJ, Ramirez AI, Garcia-Sanchez J. Immunhistokjemisk studie av menneskelig optisk nervehodet astroglia. Visjon Res. 1996;36: 2015-2028.
Turner DL, Cepko CL. En vanlig stamfar for nevroner og glia vedvarer i rotte netthinnen sent i utviklingen. Natur. 1987;328:131–136.