Gliových buněk Sítnice o Helga Kolb
Helga Kolb
Tři základní typy gliových buněk se nacházejí v lidské sítnici, Muller buněk, astroglia a mikroglie. Všechny byly pro sítnici popsány Cajalem před více než sto lety (1892).
1. Mullerovy buňky.
Mullerovy buňky jsou hlavní gliovou buňkou sítnice. Tvoří architektonické podpůrné struktury táhnoucí se radiálně přes tloušťku sítnice a jsou limity sítnice na vnější a vnitřní omezující membráně. Úplné pochopení tvaru Mullerovy buňky je nejlépe vidět po Golgiho barvení, jak ukazuje původně Cajal (1892) níže.
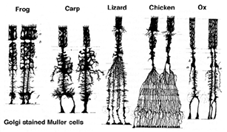
Obr. 1. Golgiho barevného Muller buňky,
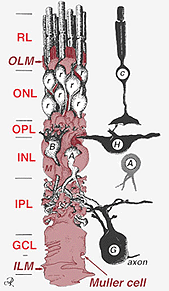
Muller buněk těla sedět ve vnitřní jaderné vrstvy a projekt nepravidelně tlusté a tenké procesy v obou směrech na vnější limitující membránou a vnitřní limitující membrány. Muller buněčných procesů zapojit mezi buňky těla neuronů v jaderné vrstvy a obálka skupin nervových procesů v plexiform vrstev (Obr. 1). Ve skutečnosti jsou sítnicové nervové procesy povoleny pouze přímým kontaktem, aniž by obklopovaly procesy Mullerových buněk, na jejich synapsích.
jediného progenitorové buňky vzniká Muller buňky a sítnice neurony(Turner a Cepko, 1987), i když zřejmě ve dvou fázích. Nejčasnější fázové neurony narozené na apikálním okraji neuroepitelu sousedící s pigmentovým epitelem produkují primární neurony sestávající z kuželových buněk,horizontálních buněk a gangliových buněk (obr. 2, vpravo). Druhá fáze buňky se také narodila na apikální rozpětí produkuje Muller buněk a rod fotoreceptory, bipolární buňky a amakrinní buňky (Reichenbach a Robinson, 1995) (Obr. 2, vlevo). Všech rozvojových neuronů a Muller buňky musí migrovat dovnitř, aby jejich konečné polohy, a to je si myslel, že Muller buněčných procesů a kmeny průvodce mnohem neuronu migrace a přímé neurite členění.
Obr. 2. 3-D schematický výkres vztah mezi Muller mobil a další sítnice neurony
uzly, které tvoří vnější limitující membrány jsou mezi Muller buňky a další Muller buněk a fotoreceptorových buněk jako robustní desmozomy nebo zonula adherens. U některých druhů gap junctions (specializované membrány sdružení a programy, které umožňují průchod malých molekul a iontů) nebo tight junctions jsou součástí těchto Muller buněčné spoje (Miller a Dowling, 1970) ale ne tak v savčích druhů, kde žádná barviva tažné kdy bylo pozorováno (Robinson et al., 1993; Reichenbach a Robinson, 1995). Povrch Muller buňky čelí pigmentového epitelu a subretinálních prostor je rozšířen o mnoho projekce Muller buněčné membrány známé jako apikální klků. Na druhé straně je vnitřní omezující membrána tvořena kuželovitou koncovkou Mullerovy buňky, ale zde nejsou vidět žádné specializované křižovatky. Mullerovy buňky také tvoří endfeet na velkých retinálních krevních cévách na vnitřním povrchu sítnice. Povrch membrány Mullerových buněk směřující ke sklivci je pokryt mukopolysacharidovým materiálem a tvoří tak skutečnou bazální membránu.
Muller buňky obsahují glykogen, mitochondrie a střední filamenta, která jsou immunoreative pro vimentinu a do jisté míry na gliový fibrilární acidicprotein (GFAP). Posledně uvedená vlákna jsou obvykle ve vnitřní polovině Mullerových buněk sítnice a jejich koncových nohou (obr. 3, vlevo), ale po traumatu sítnice, jako je oddělení sítnice, jsou vimentin i GFAP masivně upregulovány a nacházejí se v celé buňce (obr. 3, vpravo) (Guerin et al., 1990; Fisher a Lewis, 1995).
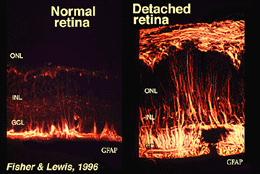
Obr. 3. GFAP imunoreaktivitu v Muller buňky,
Muller buňky mají řadu funkcí, z nichž všechny jsou životně důležité pro zdraví sítnice neurony. Mullerovy buňky fungují v symbiotickém vztahu s neurony (pro vynikající přehled viz Reichenbach a Robinson, 1995). Tak Muller mobilní funkce patří:
- Zásobujících konečného výrobku anaerobní metabolismus (rozpad glykogenu) do paliva aerobní metabolismus v nervových buňkách.
- vytírají neurální odpadní produkty, jako je oxid uhličitý a amoniak, a recyklují použité aminokyselinové vysílače.
- chrání neurony před působením přebytek neurotransmiterů, jako je glutamát pomocí dobře vyvinuté mechanismy příjmu recyklovat tento vysílač. Jsou zvláště charakterizovány přítomností vysokých koncentrací glutaminsyntázy.
- mohou se podílet jak na fagocytóze neuronálních zbytků, tak na uvolňování neuroaktivních látek, jako je GABA, taurin a dopamin.
- Oni jsou myšlenka k syntéze kyseliny retinové z retinolu (kyselina retinová je známo, že je důležité ve vývoji očí a nervového systému) (Edwards, 1994)
- kontrolují homeostázu a chrání neurony před škodlivými změny v jejich iontové prostředí tím, že se do extracelulárního K+ a k přerozdělování peněz.
- přispívají ke generaci elektroretinogram (ERG) b-vlna (Miller a Dowling, 1970; Newman a Odette, 1984), pomalý P3 součástí ERG (Karwoski a Proenza, 1977) a scotopic práh odpovědi (STR) (Frishman a Steinberg, 1989). Činí tak regulací distribuce K+ přes hranici sklivce sítnice, přes celou sítnici a lokálně ve vnitřní plexiformní vrstvě sítnice (obr. 4, od Reichenbacha a Robinsona, 1995, adaptováno od Newmana, 1989).

Obr. 4. Regulace k + Mullerovými buňkami v Mullerových buňkách
2. Astrocyty.
Astrocyty jsou gliové buňky sítnice neuroepithelium ale zadejte vyvíjející se sítnice z mozku podél rozvoj zrakového nervu (Kámen a Dreher, 1987; Chan-Ling 1994). Mají charakteristickou morfologii zploštělého buněčného těla a vláknitou řadu vyzařovacích procesů. Mezilehlá vlákna vyplňují své procesy, a tak dramaticky barví protilátkami proti GFAP(Schnitzer, 1988). Těla a procesy astrocytů jsou téměř zcela omezeny na vrstvu nervových vláken sítnice. Jejich morfologie se mění z periferie na hlavu optického nervu: ze symetrické stelátové formy v periferní sítnici (obr. 5a a b) (Schitzer, 1988)až extrémně protáhlý v blízkosti optického nervu (obr. 6 a 7).
 Obr. 5a. astrocyty v periferní sítnici. Schnitzer, 1988 |
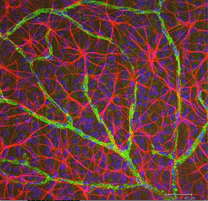 Obr. 5b. Astrocyty (červené) v sítnici potkanů mezi gangliovými buňkami (modré) a spojené s krevními cévami (zelené). S laskavým svolením Laura Fernandez-Nicolas Sanchez a Cuenca |
V imunocytochemické barvení (Obr. 5b) a při intracelulárních injekcích HRP (obr. 7) obarvené astrocyty jasně vykazují procesy zarovnané podél axonů gangliových buněk, které procházejí vrstvou nervových vláken. V distribuci, astrocyty dosáhnout svého vrcholu na hlavou optického nervu a mají poměrně rovnoměrný pokles hustoty v vyzařující kroužky z nervu. Nejsou přítomny v avaskulární fovea nebo ora serrata.
 Obr. 6. Astrocyty v centrální sítnici. Schnitzer, 1988 |
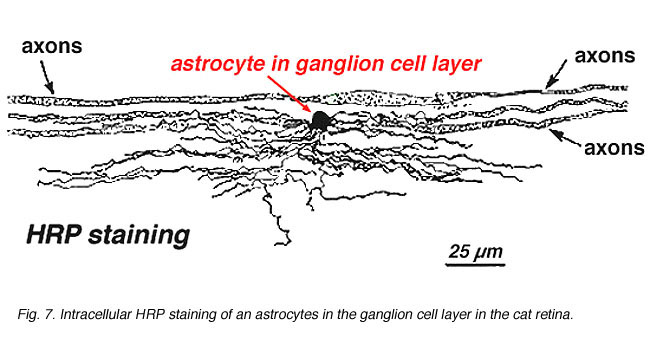 Obr. 7. Astrocyty v kočičí sítnici obarvené intracelulární injekcí HRP. S laskavým svolením Ralpha Nelsona. |
Tlusté a tenké astrocyty byly odlišeny na základě barvení s protilátkami proti GFAP (Trevino et al., 1996). Tak astrocyty jsou uspořádány na povrchu gangliové buňky axon svazky jako oni samozřejmě do zrakového nervu tvoří trubky, jehož prostřednictvím axonů běh (Obr. 8). Gap junctions a zonula adherens junctions byly popsány mezi astrocytic procesy v kočičí sítnice (Höllander et al., 1991).
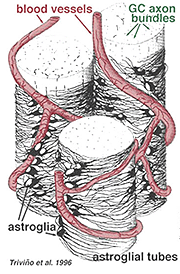
Obr. 8. 3-D blok astrocyty uspořádány na povrchu gangliové buňky axon svazky
krevní cévy, chod a mezi gangliových buněk svazky jsou také zahrnuty do obou procesů, a dokonce i příležitostné mobilní tělo z astrocytu. Funkce astrocytů obalující axony gangliových buněk a vztah k krevních cév, nervových vláken vrstva naznačuje, že jsou axonální a cévní nervové pochvy a část krve-bariéra mozku. Podobně jako Mullerovy buňky, je známo, že obsahují hojný glykogen a mohou tvořit výživnou službu při poskytování glukózy neuronům. Kromě toho pravděpodobně hrají roli v iontové homeostáze při regulaci extracelulárních hladin draslíku a metabolismu neurotransmiterů, jako je GABA.
3. Mikrogliální buňky.
třetí typ gliových buněk je údajně mezodermálního původu, a proto striktně řečeno nejsou neurogliální, stejně jako astrocyty a Mullerovy buňky. Vstupují do sítnice shodně s mezenchymálními prekurzory retinálních krevních cév ve vývoji (Chan-Ling, 1994). Mikrogliální buňky jsou všudypřítomné v lidské sítnici, která se nachází v každé vrstvě sítnice.
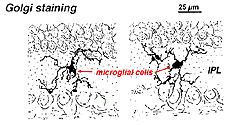
V Golgiho-barevné sítnice vypadají jako podivné, multipolární podoby s malými buněk těla a nepravidelné krátké procesy. Ve skutečnosti se v přípravcích Golgiho někdy mýlili s nervovými buňkami, zejména když leží v jaderné vrstvě s jedinou orientací svých procesů v plexiformní vrstvě.
Obr. 9. Golgiho barvení mikrogliálních buněk
mikrogliální buňky mohou být dvou typů. Předpokládá se, že jedna forma vstupuje do sítnice v raných stádiích vývoje z mezenchymu zrakového nervu a po většinu života sítnice spí ve vrstvách sítnice. Jiná forma mikroglie se zdají mít krevní buňky, možné pocházejících z plavidla pericyty (Bojkot a Hopkins, 1981; Gallego, 1986). Oba typy mohou být stimulovány do makrofagické funkce po traumatu sítnice a poté se zapojují do fagocytózy degenerujících neuronů sítnice.

Obr. 10a. Mikrogliální buňka barvená lektinem z Chan-Lin, 1994.
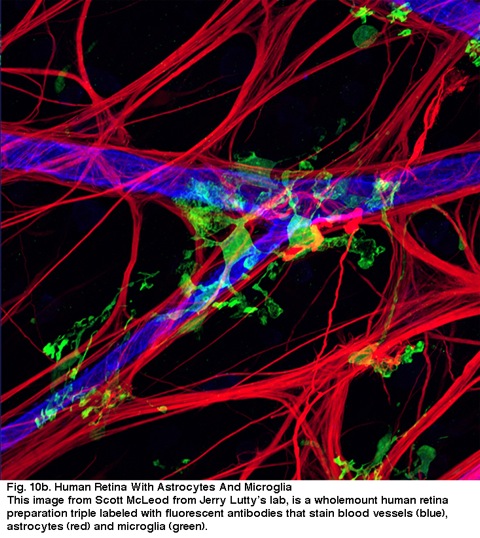
Obr. 10b. Lidská sítnice s Astrocyty a Mikroglie. Tento snímek od Scotta Mcleoda od Jerryho Lutty laboratoři, je wholemount lidské sítnice příprava triple laneled s fluorescenční protilátky, které skvrna cévy (modrá), astrocyty (červená) a mikroglie (zelená).
4. Odkaz.
bojkot BB, Hopkins JM. Mikroglie v sítnici opic a jiných savců; jeho odlišnost od jiných typů glia a horizontálních buněk. Neurověda.1981;6:679–688.
Cajal SR. In: Thorpe SA, Glickstein M, translators. 1892. Struktura sítnice. Springfield (IL): Thomas; 1972.
Chan-Ling T. gliální, neuronální a vaskulární interakce v savčí sítnici. Pořada. Ret. Eye Res. 1994; 13: 357-389.
Edwards RB. Biosyntéza kyseliny retinové müllerovými gliovými buňkami: model pro centrální nervový systém? Pořada. Ret. Eye Res. 1994; 13: 231-242.
Fisher SK, Lewis GP. Fotoreceptory a dále: buněčné a molekulární účinky odchlípení sítnice.2nd Great Basin Visual Science Symposium, II, University of Utah Press. 1995
Frishman LJ, Steinberg RH. Světlo evokované zvýšení o v proximální části sítnice kočky přizpůsobené tmě. J Neurofyziol. 1989;61:1233–1243.
Gallego a. srovnávací studie o horizontálních buňkách a poznámka o mikrogliálních buňkách. Pořada. Ret. Res.1986;5:165-206.
Guerin CJ, Anderson DH, Fisher SK. Ke změnám v imunoznačení mezilehlých vláken dochází v reakci na oddělení sítnice a opětovné připojení u primátů.Investovat. Oční. Visum. Věda. 1990;31:1474–1482.
Karwoski CJ, Proenza LM. Vztah mezi odpovědí Mullerových buněk, lokálním transretinálním potenciálem a tokem draslíku. J Neurofyziol. 1977;40:244–259.
Miller RF, Dowling JE. Intracelulární odpovědi Muller (gliových) buněk z mudpuppy sítnice: jejich vztah k b-vlna elektroretinogram. J Neurofyziol.1970;33:323–341.
Newman EA, Odette LL. Model generování elektroretinogramu b-vlny: test hypotézy k+. J Neurofyziol. 1984;51:164–182.
Newman EA. Elektrofyziologie gliových buněk sítnice. Pořada. Ret. Res.1989;8:153-172.
Reichenbach A, Robinson SR. Zapojení Müllerových buněk do vnější sítnice. V: Djamgoz MBA, Archer SN, Vallerga S, editoři. Neurobiologie a klinické aspekty vnější sítnice. London: Chapman & Hall; 1995. s. 395-416.
Robinson SR, Hampson ECM, Munro MN, Vaney DI. Jednosměrná vazba mezer mezi neurogliemi. Proc. Austr. Neurovědci. SOC. 1993;3:167.
Schnitzer J. astrocyty v savčí sítnici. Pořada. Ret. Res.1988;7:209-232.
kámen J, Makarov F, Hollander h. gliální ensheathment soma a axon hillock retinálních gangliových buněk. Návštěvník. 12:273–279.
Stone J, Dreher z. vztah mezi astrocyty, gangliovými buňkami a vaskulaturou sítnice. J Comp Neurol. 1987;255:35–49.
Trivino A, Ramirez JM, Salazar JJ, Ramirez AI, Garcia-Sanchez J. Imunohistochemická studie astroglie hlavy lidského optického nervu. Vision Res. 1996; 36: 2015-2028.
Turner DL, Cepko CL. Společný progenitor neuronů a glie přetrvává v sítnici potkanů v pozdním vývoji. Povaha. 1987;328:131–136.