komórki glejowe siatkówki by Helga Kolb
Helga Kolb
trzy podstawowe typy komórek glejowych znajdują się w ludzkiej siatkówce, komórkach Mullera, astroglii i mikrogleju. Wszystkie zostały opisane dla siatkówki przez Cajala ponad sto lat temu (1892).
1. Komórki Mullera.
komórki Mullera są główną komórką glejową siatkówki. Tworzą architektoniczne struktury nośne rozciągające się promieniowo na całej grubości siatkówki i są granicami siatkówki odpowiednio na zewnętrznej i wewnętrznej błonie ograniczającej. Pełne zrozumienie kształtu komórki Mullera jest najlepiej widoczne po barwieniu Golgiego, jak pokazano pierwotnie przez Cajala (1892) poniżej.
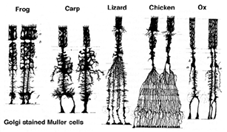
rys. 1. Komórki Golgiego
ciała komórek Mullera siedzą w wewnętrznej warstwie jądrowej i projektują nieregularnie grube i cienkie procesy w obu kierunkach do zewnętrznej błony ograniczającej i do wewnętrznej błony ograniczającej. Procesy komórkowe Mullera insynuują się między ciałami komórkowymi neuronów w warstwach jądrowych a grupami obwiedniowymi procesów neuronowych w warstwach plexi (Fig. 1). W rzeczywistości procesy nerwowe siatkówki są dozwolone tylko w bezpośrednim kontakcie, bez otaczania procesów komórkowych Mullera, w ich synapsach.
pojedyncza komórka progenitorowa powoduje powstanie zarówno komórek Mullera, jak i neuronów siatkówki(Turner and Cepko, 1987), chociaż najwyraźniej w dwóch fazach. Najwcześniejsze neurony fazowe urodzone na wierzchołkowym brzegu neuroepitelium przylegającym do nabłonka pigmentowego wytwarzają neurony pierwotne składające się z komórek stożkowych, komórek poziomych i komórek zwojowych (Fig. 2, prawo). Druga faza komórek również urodzonych na brzegach wierzchołkowych wytwarza komórki Mullera i fotoreceptory rod, komórki dwubiegunowe i komórki amakryny (Reichenbach i Robinson, 1995) (Fig. 2, po lewej). Wszystkie rozwijające się neurony i komórki Mullera muszą migrować do środka do swojej ostatecznej pozycji i uważa się, że procesy komórkowe i pnie Mullera kierują większością migracji neuronów i kierują różnicowaniem neurytu.
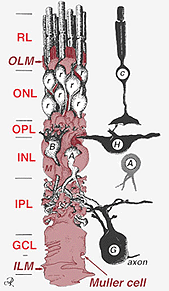
rys. 2. 3-D schematyczny rysunek zależności między komórką Mullera a innymi neuronami siatkówki
połączenia tworzące zewnętrzną błonę ograniczającą znajdują się między komórkami Mullera a innymi komórkami Mullera i komórkami fotoreceptorowymi jako wytrzymałe desmosomy lub przylegające zonula. W niektórych gatunkach połączenia szczelinowe (wyspecjalizowane związki membranowe i kanały, które umożliwiają przejście małych cząsteczek i jonów) lub szczelne połączenia są częścią tych połączeń komórkowych Mullera (Miller and Dowling, 1970), ale nie tak u gatunków ssaków, w których nigdy nie zaobserwowano sprzężenia barwnika (Robinson i wsp., 1993; Reichenbach i Robinson, 1995). Powierzchnia komórki Mullera skierowana do nabłonka pigmentowego i przestrzeni podretinalnej jest rozszerzona przez wiele projekcji błony komórkowej Mullera znanej jako kosmki wierzchołkowe. Wewnętrzna membrana ograniczająca, z drugiej strony, jest utworzona przez stożkową końcówkę komórki Mullera, ale nie widać tu wyspecjalizowanych połączeń. Komórki Mullera tworzą również stopy końcowe na dużych naczyniach krwionośnych siatkówki na wewnętrznej powierzchni siatkówki. Powierzchnia błony komórkowej Mullera zwrócona do ciała szklistego pokryta jest materiałem mukopolisacharydowym i w ten sposób tworzy prawdziwą błonę podstawną.
komórki Mullera zawierają glikogen, mitochondria i włókna pośrednie, które są immunoreatywne dla wimentiny i w pewnym stopniu dla glejowej włóknistej kwaśnej proteiny (GFAP). Te ostatnie włókna znajdują się zwykle w wewnętrznej połowie siatkówkowych komórek Mullera i ich stopie końcowym (Fig. 3, po lewej), ale po urazie siatkówki, takim jak odwarstwienie siatkówki, zarówno wimentin, jak i GFAP są masowo regulowane i znajdują się w całej komórce (rys. 3, prawo) (Guerin et al., 1990; Fisher and Lewis, 1995).
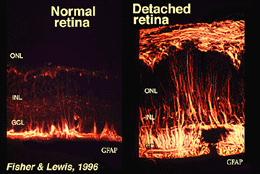
rys. 3. GFAP immunoreaktywność w komórkach Mullera
komórki Mullera mają szereg funkcji, z których wszystkie są niezbędne dla zdrowia neuronów siatkówki. Komórki Mullera funkcjonują w symbiotycznej relacji z neuronami (dla doskonałego przeglądu patrz Reichenbach and Robinson, 1995). Zatem funkcje komórek Mullera obejmują:
- dostarczanie końcowych produktów metabolizmu beztlenowego (rozpad glikogenu) do napędzania metabolizmu tlenowego w komórkach nerwowych.
- usuwają neuronowe produkty odpadowe, takie jak dwutlenek węgla i amoniak, i przetwarzają zużyte nadajniki aminokwasów.
- chronią neurony przed ekspozycją na nadmiar neuroprzekaźników, takich jak glutaminian, wykorzystując dobrze rozwinięte mechanizmy wychwytu do recyklingu tego nadajnika. Szczególnie charakteryzują się obecnością wysokich stężeń syntazy glutaminowej.
- mogą być zaangażowane zarówno w fagocytozę szczątków neuronalnych, jak i uwalnianie substancji neuroaktywnych, takich jak GABA, tauryna i dopamina.
- uważa się, że syntetyzują kwas retinowy z retinolu (kwas retinowy jest znany jako ważny w rozwoju oka i układu nerwowego) (Edwards, 1994)
- kontrolują homeostazę i chronią neurony przed szkodliwymi zmianami w ich środowisku jonowym poprzez pobieranie zewnątrzkomórkowego K+ i redystrybucję go.
- przyczyniają się do generowania fali B elektroretinogramu (Erg) (Miller i Dowling, 1970; Newman i Odette, 1984), powolnego komponentu P3 ERG (Karwoski i Proenza, 1977) i scotopic threshold response (STR) (Frishman i Steinberg, 1989). Robią to poprzez regulację rozkładu K+ na granicy ciała szklistego siatkówki, na całej siatkówce i lokalnie w wewnętrznej warstwie splotu siatkówki (Fig. 4, z Reichenbach i Robinson, 1995, adaptacja z Newman, 1989).

rys. 4. Regulacja K+ przez komórki Mullera w komórkach Mullera
2. Astrocyty.
astrocyty nie są komórkami glejowymi neuroepitelium siatkówki, ale wchodzą do rozwijającej się siatkówki z mózgu wzdłuż rozwijającego się nerwu wzrokowego (Stone i Dreher, 1987; Chan-Ling 1994). Charakteryzują się charakterystyczną morfologią spłaszczonego ciała komórkowego i włóknistym szeregiem procesów promieniujących. Włókna pośrednie wypełniają swoje procesy i w ten sposób dramatycznie plamią przeciwciałami przeciwko GFAP (Schnitzer, 1988). Astrocyty ciała komórkowe i procesy są prawie całkowicie ograniczone do warstwy włókien nerwowych siatkówki. Ich morfologia zmienia się od obrzeża do głowy nerwu wzrokowego: od symetrycznej postaci kostnej w siatkówce obwodowej (Fig. 5a i b) (Schitzer, 1988) do bardzo wydłużonego w pobliżu nerwu wzrokowego (rys. 6 i 7).
 rys. 5A. astrocyty w siatkówce obwodowej. Schnitzer, 1988 |
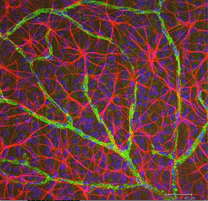 rys. 5b. Astrocyty (czerwone) w siatkówce szczura wśród komórek zwojowych (niebieskie) i związane z naczyniami krwionośnymi (zielone). Dzięki uprzejmości Laury Fernandez-Sanchez i Nicolasa Cuenca |
w barwieniu immunocytochemicznym (rys. 5b) oraz w zastrzykach wewnątrzkomórkowych HRP (ryc. 7) barwione astrocyty wyraźnie wykazują procesy wyrównane wzdłuż aksonów komórek zwojowych przebiegających przez warstwę włókien nerwowych. W dystrybucji astrocyty osiągają swój szczyt na głowie nerwu wzrokowego i mają dość równomierny spadek gęstości w promieniujących pierścieniach z głowy nerwu. Nie występują w jajowodzie ani w ora serrata.
 rys. 6. Astrocyty w środkowej siatkówce. Schnitzer, 1988 |
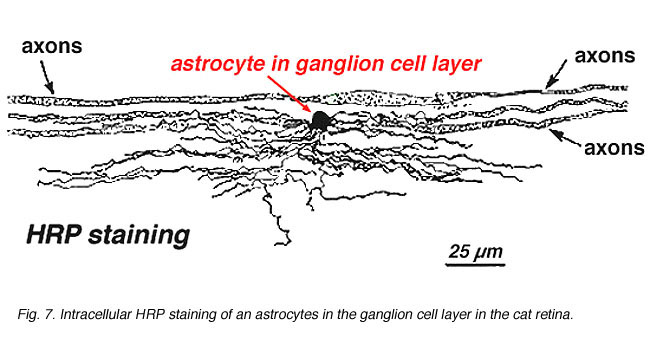 rys. 7. Astrocyty w siatkówce kotów zabarwione przez wewnątrzkomórkowe wstrzyknięcie HRP. Dzięki uprzejmości Ralpha Nelsona. |
grube i cienkie astrocyty zostały wyróżnione na podstawie barwienia przeciwciałami przeciwko GFAP (Trevino et al., 1996). W ten sposób astrocyty są rozmieszczone na powierzchni wiązek aksonów komórek zwojowych, gdy przebiegają do głowy nerwu wzrokowego, tworząc rurkę, przez którą biegną aksony (Fig. 8). Węzły szczelinowe i węzły zonula adherens zostały opisane między procesami astrocytowymi w siatkówce kota (Höllander et al., 1991).
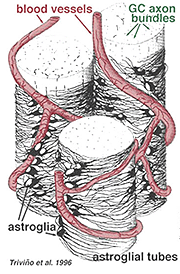
rys. 8. 3 – D Blok astrocytów ułożonych na powierzchni wiązek aksonów komórek zwojowych
naczynia krwionośne biegnące w i między wiązkami komórek zwojowych są również objęte przez oba procesy, a nawet sporadyczne ciało komórkowe astrocytów. Funkcja astrocytów otaczających aksony komórek zwojowych i związek z naczyniami krwionośnymi warstwy włókien nerwowych sugeruje, że są to aksonalne i naczyniowe osłonki glejowe i stanowią część bariery krew-mózg. Podobnie jak komórki Mullera, wiadomo, że zawierają obfity glikogen i mogą tworzyć odżywczą usługę dostarczania glukozy do neuronów. Ponadto prawdopodobnie pełnią rolę w homeostazie jonowej w regulacji pozakomórkowego poziomu potasu i metabolizmu neuroprzekaźników, takich jak GABA.
3. Komórki mikroglialne.
trzeci typ komórek glejowych jest podobno pochodzenia mezodermalnego, a zatem, ściśle mówiąc, nie są neuroglialne, podobnie jak astrocyty i komórki Mullera. Dostają się do siatkówki wraz z mezenchymalnymi prekursorami naczyń krwionośnych siatkówki w rozwoju (Chan-Ling, 1994). Komórki mikroglialne są wszechobecne w ludzkiej siatkówce, które znajdują się w każdej warstwie siatkówki.
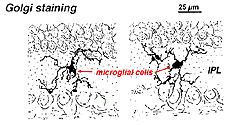
w siatkówce Golgiego wyglądają jak dziwne, wielobiegunowe formy z małymi ciałami komórkowymi i nieregularnymi krótkimi procesami. W preparatach Golgiego czasami mylono je z komórkami nerwowymi, zwłaszcza gdy leżą w warstwie jądrowej z pojedynczą orientacją ich procesów w warstwie plexi.
rys. 9. Golgiego barwienie komórek mikroglialnych
komórki Mikroglialne mogą być dwóch typów. Uważa się, że jedna forma wchodzi do siatkówki we wczesnych stadiach rozwoju z mezenchymy nerwu wzrokowego i leży uśpiona w warstwach siatkówki przez większą część życia siatkówki. Inną formą mikrogleju wydają się być komórki krwiopochodne, możliwe pochodzące z pericytów naczyń (Boycott and Hopkins, 1981; Gallego, 1986). Oba typy mogą być stymulowane do funkcji makrofagicznej po urazie siatkówki, a następnie angażują się w fagocytozę zwyrodniałych neuronów siatkówki.

rys. 10A. Lectin-stained microglial cell from Chan-Lin, 1994.
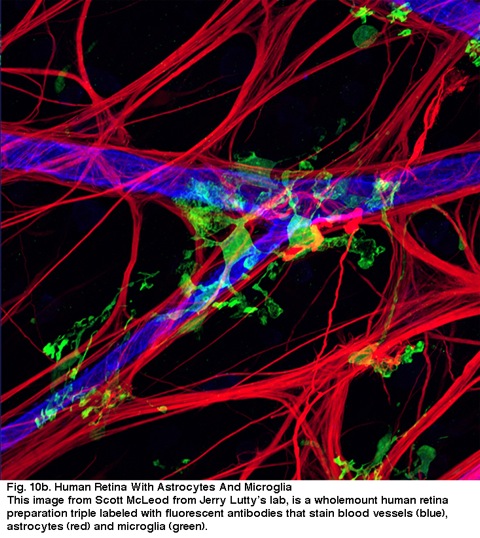
10B. ludzka siatkówka z astrocytami i Mikroglejem. Ten obraz z Scott McLeod z laboratorium Jerry 'ego Lutty’ ego, jest wholemount preparat ludzkiej siatkówki potrójnie laneled z przeciwciałami fluorescencyjnymi, które plamią naczynia krwionośne (niebieski), astrocyty (czerwony) i mikrogleju (zielony).
4. Referencje.
bojkot BB, Hopkins JM. Mikroglej w siatkówce małp i innych ssaków; jego odróżnienie od innych rodzajów gleju i komórek poziomych. Neurobiologia.1981;6:679–688.
Cajal SR. w: Thorpe SA, Glickstein M, Tłumacze. 1892. Struktura siatkówki. Springfield (IL): Thomas; 1972.
Chan-Ling T. interakcje glejowe, neuronalne i naczyniowe w siatkówce ssaków. Prog. Ret. Eye Res. 1994;13: 357-389.
Edwards RB. Biosynteza kwasu retinowego przez komórki glejowe Müllera: model dla ośrodkowego układu nerwowego? Prog. Ret. Eye Res. 1994;13:231-242.
Fisher SK, Lewis GP. Fotoreceptory i nie tylko: komórkowe i molekularne efekty odwarstwienia siatkówki.2nd Great Basin Visual Science Symposium, II, University of Utah Press. 1995
Frishman LJ, Steinberg Rh. Wywołane światłem wzrosty o w proksymalnej części ciemnej siatkówki kota. J Neurofizjol. 1989;61:1233–1243.
Gallego A. Comparative studies on horizontal cells and a note on microglial cells. Prog. Ret. Res. 1986; 5:165-206.
Guerin CJ, Anderson DH, Fisher SK. Zmiany w immunolabelingu włókien pośrednich występują w odpowiedzi na odwarstwienie siatkówki i ponowne przywarstwienie u naczelnych.Inwestuj. Ophthal. Vis. Sci. 1990;31:1474–1482.
Karwoski CJ, Proenza LM. Związek między odpowiedziami komórek Mullera, lokalnym potencjałem przezskórnym i strumieniem potasu. J Neurofizjol. 1977;40:244–259.
Miller RF, Dowling JE. Odpowiedzi wewnątrzkomórkowe komórek Mullera (glejowych) siatkówki mudpuppy ’ ego: ich związek z falą B elektroretinogramu. J Neurofizjol.1970;33:323–341.
Newman EA, Odette LL. Model generacji fali B elektroretinogramu: test hipotezy K+. J Neurofizjol. 1984;51:164–182.
Newman EA. Elektrofizjologia komórek glejowych siatkówki. Prog. Ret. Res. 1989; 8:153-172.
Reichenbach a, Robinson SR. Udział komórek Müllera w zewnętrznej siatkówce. In: Djamgoz MBA, Archer SN, Vallerga S, editors. Neurobiologia i kliniczne aspekty siatkówki zewnętrznej. London: Chapman & Hall; 1995. S. 395-416.
Robinson SR, Hampson ECGM, Munro MN, Vaney DI. Jednokierunkowe sprzężenie szczelin między neuroglią. Proc. Austr. Neurosci. Soc. 1993;3:167.
Schnitzer J. Astrocytes in mammalian retina. Prog. Ret. Res. 1988; 7:209-232.
Stone J, Makarov F, Hollander H. the glejal ensheathment of the soma and axon hillock of retinal ganglion cells. Vis Neurosci. 12:273–279.
Stone J, Dreher Z. związek między astrocytami, komórkami zwojowymi i unaczynieniem siatkówki. J Comp Neurol. 1987;255:35–49.
Trivino a, Ramirez JM, Salazar JJ, Ramirez AI, Garcia-Sanchez J. Immunohistochemical study of human optic nerve head astroglia. Vision Res. 1996;36: 2015-2028.
Turner DL, Cepko CL. Wspólny progenitor dla neuronów i gleju utrzymuje się w siatkówce szczurów pod koniec rozwoju. Natura. 1987;328:131–136.