celulele gliale ale retinei de Helga Kolb
Helga Kolb
trei tipuri de bază de celule gliale se găsesc în retina umană, celulele Muller, astroglia și microglia. Toate au fost descrise pentru retină de Cajal acum mai bine de o sută de ani (1892).
1. Celulele Muller.celulele Muller sunt principala celulă glială a retinei. Ele formează structuri de sprijin arhitectural care se întind radial pe grosimea retinei și sunt limitele retinei la membrana limitativă exterioară și, respectiv, interioară. O înțelegere completă a formei unei celule Muller este cel mai bine văzută după colorarea Golgi, așa cum se arată inițial de Cajal (1892) de mai jos.
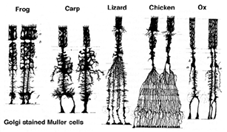
Fig. 1. Celulele Muller colorate Golgi
corpurile celulelor Muller stau în stratul nuclear interior și proiectează procese neregulate groase și subțiri în ambele direcții către membrana limitativă exterioară și către membrana limitativă interioară. Procesele celulare Muller se insinuează între corpurile celulare ale neuronilor din straturile nucleare și grupurile de plicuri ale proceselor neuronale din straturile plexiforme (Fig. 1). De fapt, procesele neuronale retiniene sunt permise numai contactul direct, fără a înfășura procesele celulare Muller, la sinapsele lor.
o singură celulă progenitoare dă naștere atât celulelor Muller, cât și neuronilor retinieni(Turner și Cepko, 1987), deși aparent în două faze. Cei mai vechi neuroni de fază născuți la marginea apicală a neuroepiteliului adiacent epiteliului pigmentar produc neuroni primari constând din celule conice, celule orizontale și celule ganglionare (Fig. 2, dreapta). A doua fază a celulelor născute și la marginile apicale produce celule Muller și fotoreceptori de tijă, celule bipolare și celule amacrine (Reichenbach și Robinson, 1995) (Fig. 2, stânga). Toți neuronii în curs de dezvoltare și celulele Muller trebuie să migreze spre interior în poziția lor finală și se crede că procesele și trunchiurile celulelor Muller ghidează o mare parte din migrațiile neuronilor și direcționează diferențierile neurite.
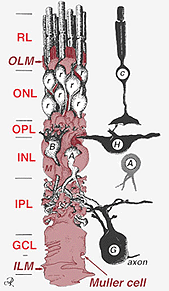
Fig. 2. Desen schematic 3-D al relației dintre celula Muller și alți neuroni retinieni
joncțiunile care formează membrana limitativă exterioară se află între celulele Muller și alte celule Muller și celulele fotoreceptoare ca desmosomi rezistenți sau zonula adherens. La unele specii joncțiunile gap (asociații și canale de membrană specializate care permit trecerea moleculelor mici și a ionilor) sau joncțiunile strânse fac parte din aceste joncțiuni celulare Muller (Miller și Dowling, 1970), dar nu la speciile de mamifere unde nu s-a observat niciodată cuplarea colorantului (Robinson și colab., 1993; Reichenbach și Robinson, 1995). Suprafața celulei Muller cu care se confruntă epiteliul pigmentar și spațiul subretinal este extinsă de multe proiecții ale membranei celulare Muller cunoscute sub numele de vilozități apicale. Membrana limitativă interioară, pe de altă parte, este formată din picioarele conice ale celulei Muller, dar aici nu se văd joncțiuni specializate. Celulele Muller formează, de asemenea, picioarele finale pe vasele mari de sânge retiniene de la suprafața interioară a retinei. Suprafața membranei celulare Muller orientată spre vitros este acoperită cu un material mucopolizaharidic și astfel formează o adevărată membrană bazală.
celulele Muller conțin glicogen, mitocondrii și filamente intermediare care sunt imunoreative pentru vimentină și într-o oarecare măsură pentru acid fibrilar glialproteină (GFAP). Ultimele filamente sunt în mod normal în jumătatea interioară a celulelor Muller retiniene și a picioarelor lor finale (Fig. 3, stânga), dar în urma traumatismelor retinei, cum ar fi detașarea retinei, atât vimentina, cât și GFAP sunt reglate masiv și se găsesc în toată celula (Fig. 3, dreapta) (Guerin și colab., 1990; Fisher și Lewis, 1995).
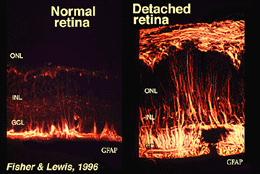
Fig. 3. Imunoreactivitatea GFAP în celulele Muller
celulele Muller au o serie de funcții, toate fiind vitale pentru sănătatea neuronilor retinieni. Celulele Muller funcționează într-o relație simbiotică cu neuronii (pentru o revizuire excelentă vezi Reichenbach și Robinson, 1995). Astfel, funcțiile celulelor Muller includ:
- furnizarea de produse finale ale metabolismului anaerob (descompunerea glicogenului) pentru a alimenta metabolismul aerob în celulele nervoase.
- elimină deșeurile neuronale, cum ar fi dioxidul de carbon și amoniacul și reciclează emițătoarele de aminoacizi uzate.
- protejează neuronii de expunerea la neurotransmițători în exces, cum ar fi glutamatul, folosind mecanisme de absorbție bine dezvoltate pentru a recicla acest transmițător. Acestea se caracterizează în special prin prezența unor concentrații ridicate de glutamină sintază.
- ele pot fi implicate atât în fagocitoza resturilor neuronale, cât și în eliberarea substanțelor neuroactive, cum ar fi GABA, taurina și dopamina.
- se crede că sintetizează acidul retinoic din retinol (acidul retinoic este cunoscut a fi important în dezvoltarea ochiului și a sistemului nervos) (Edwards, 1994)
- controlează homeostazia și protejează neuronii de schimbările dăunătoare din mediul lor ionic prin preluarea K+ extracelular și redistribuirea acestuia.
- ele contribuie la generarea electroretinogramei (ERG) B-wave (Miller și Dowling, 1970; Newman și Odette, 1984), componenta lentă P3 a ERG (Karwoski și Proenza, 1977) și răspunsul pragului scotopic (STR) (Frishman și Steinberg, 1989). Acestea fac acest lucru prin reglarea distribuției K+ pe marginea vitroasă a retinei, pe întreaga retină și local în stratul plexiform interior al retinei (Fig. 4, din Reichenbach și Robinson, 1995, adaptat din Newman, 1989).

Fig. 4. Reglarea K + de către celulele Muller în celulele Muller
2. Astrocite.
astrocitele nu sunt celule gliale ale neuroepiteliului retinian, ci intră în retina în curs de dezvoltare din creier de-a lungul nervului optic în curs de dezvoltare (Stone și Dreher, 1987; Chan-Ling 1994). Ele au o morfologie caracteristică a unui corp celular aplatizat și o serie fibroasă de procese radiante. Filamentele intermediare își umplu procesele și astfel se colorează dramatic cu anticorpi împotriva GFAP (Schnitzer, 1988). Corpurile și procesele celulelor astrocite sunt aproape în întregime limitate la stratul de fibre nervoase al retinei. Morfologia lor se schimbă de la periferie la capul nervului optic: de la o formă stelată simetrică în retina periferică (Fig. 5a și b) (Schitzer, 1988) la extrem de alungit în apropierea nervului optic (Fig. 6 și 7).
 Fig. 5a. astrocite în retina periferică. Schnitzer, 1988 |
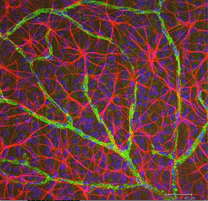 Fig. 5b. Astrocite (roșu) în retina șobolanului printre celulele ganglionare (albastru) și asociate cu vasele de sânge (verde). Prin amabilitatea Laurei Fernandez-Sanchez și Nicolas Cuenca |
în colorarea imunocitochimică (Fig. 5b) și în injecții intracelulare HRP (Fig. 7) astrocitele colorate prezintă în mod clar procese aliniate de-a lungul axonilor celulelor ganglionare care curg prin stratul de fibre nervoase. În distribuție, astrocitele ating vârful pe capul nervului optic și au o scădere destul de uniformă a densității în inelele radiante din capul nervului. Nu sunt prezente în foveea avasculară sau ora serrata.
 Fig. 6. Astrocite în retina centrală. Schnitzer, 1988 |
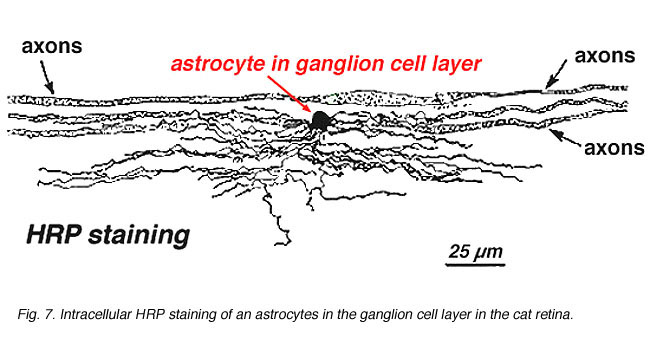 Fig. 7. Astrocite în retina pisicii colorate prin injectarea intracelulară a HRP. Prin amabilitatea lui Ralph Nelson. |
astrocitele groase și subțiri au fost distinse pe baza colorării cu anticorpi la GFAP (Trevino și colab., 1996). Astfel, astrocitele sunt dispuse pe suprafața mănunchiurilor axonice ale celulelor ganglionare în timp ce curg în capul nervului optic formând un tub prin care trec axonii (Fig. 8). Joncțiunile Gap și joncțiunile zonula adherens au fost descrise între procesele astrocitice din retina pisicii (h Elixtllander și colab., 1991).
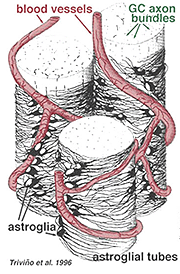
Fig. 8. Bloc 3-D de astrocite dispuse pe suprafața mănunchiurilor axonice ale celulelor ganglionare
vasele de sânge care circulă în și printre mănunchiurile celulare ganglionare sunt, de asemenea, acoperite de ambele procese și chiar de un corp celular ocazional al unui astrocite. Funcția astrocitelor care înconjoară axonii celulelor ganglionare și relația cu vasele de sânge ale stratului de fibre nervoase sugerează că acestea sunt învelișuri gliale axonale și vasculare și fac parte dintr-o barieră hematoencefalică. Similar cu celulele Muller, se știe că conțin glicogen abundent și pot forma un serviciu nutritiv în furnizarea de glucoză neuronilor. În plus, ele servesc probabil un rol în homeostazia Ionică în reglarea nivelurilor extracelulare de potasiu și metabolismul neurotransmițătorilor precum GABA.
3. Celule microgliale.
al treilea tip de celule gliale se presupune că este de origine mezodermică și, prin urmare, strict vorbind nu sunt neurogliale la fel ca astrocitele și celulele Muller. Acestea intră în retină coincident cu precursorii mezenchimali ai vaselor de sânge retiniene în dezvoltare (Chan-Ling, 1994). Celulele microgliale sunt omniprezente în retina umană, fiind găsite în fiecare strat al retinei.
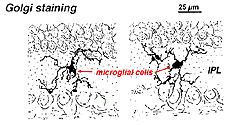
în retina colorată de Golgi arată ca forme ciudate, multipolare, cu corpuri celulare mici și procese scurte neregulate. De fapt, în preparatele Golgi au fost uneori confundate cu celulele nervoase, în special atunci când se află într-un strat nuclear cu o singură orientare a proceselor lor în stratul plexiform.
Fig. 9. Colorarea Golgi a celulelor microgliale
celulele microgliale pot fi de două tipuri. Se crede că o formă intră în retină în stadiile incipiente de dezvoltare din mezenchimul nervului optic și se află latentă în straturile retiniene pentru o mare parte din viața retinei. Cealaltă formă de microglie pare a fi celulele transmise de sânge, posibil provenind din pericitele vaselor (boicot și Hopkins, 1981; Gallego, 1986). Ambele tipuri pot fi stimulate într-o funcție macrofagică după trauma retinei și apoi se angajează în fagocitoza neuronilor retinieni degenerați.

Fig. 10a. Celulă microglială colorată cu lectină din Chan-Lin, 1994.
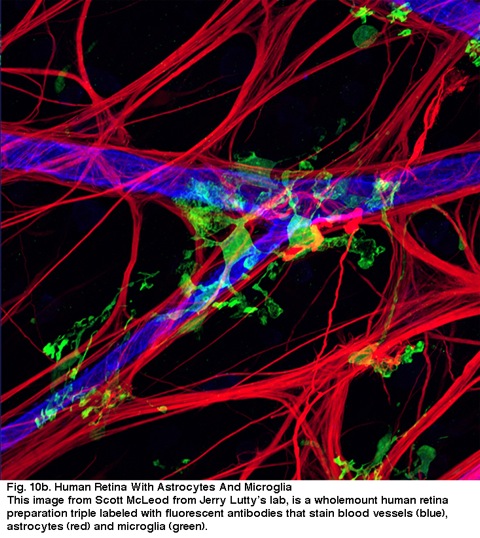
Fig. 10B. retina umană cu astrocite și microglii. Această imagine de la Scott McLeod de la laboratorul lui Jerry Lutty este o pregătire integrală a retinei umane triplă cu anticorpi fluorescenți care pătează vasele de sânge (albastru), astrocite (roșu) și microglia (verde).
4. Referințe.
boicot BB, Hopkins JM. Microglia în retina maimuței și a altor mamifere; distincția sa față de alte tipuri de glia și celule orizontale. Neuroștiințe.1981;6:679–688.
Cajal SR. în: Thorpe SA, Glickstein M, traducători. 1892. Structura retinei. Springfield( IL): Thomas; 1972.Chan-Ling T. interacțiuni gliale, neuronale și vasculare în retina mamiferelor. Prog. Ret. Eye Res. 1994; 13: 357-389.
Edwards RB. Biosinteza acidului retinoic de către celulele gliale m Inkller: un model pentru sistemul nervos central? Prog. Ret. Eye Res. 1994; 13: 231-242.
Fisher SK, Lewis GP. Fotoreceptori și nu numai: efecte celulare și moleculare ale detașării retinei.Al 2-lea Simpozion de științe vizuale din Marele Bazin, ii, Universitatea din Utah Press. 1995
Frishman LJ, Steinberg RH. Creșteri evocate de lumină în o în porțiunea proximală a retinei pisicii adaptate la întuneric. J Neurofiziol. 1989;61:1233–1243. Gallego A. studii Comparative asupra celulelor orizontale și o notă asupra celulelor microgliale. Prog. Ret. Res. 1986; 5: 165-206.
Guerin CJ, Anderson DH, Fisher SK. Modificările imunomodulării filamentului intermediar apar ca răspuns la detașarea retinei și reatașarea la primate.Investește. Ophal. Vis. Sci. 1990;31:1474–1482.
Karwoski CJ, Proenza LM. Relația dintre răspunsurile celulelor Muller, un potențial transretinal local și fluxul de potasiu. J Neurofiziol. 1977;40:244–259.
Miller RF, Dowling JE. Răspunsurile intracelulare ale celulelor Muller (gliale) ale retinei mudpuppy: relația lor cu valul B al electroretinogramei. J Neurofiziol.1970;33:323–341.
Newman EA, Odette II. Modelul generării electroretinogramei B-wave: un test al ipotezei K+. J Neurofiziol. 1984;51:164–182.
Newman EA. Electrofiziologia celulelor gliale retiniene. Prog. Ret. Res. 1989; 8: 153-172.
Reichenbach A, Robinson SR. Implicarea celulelor m-Xicller în retina exterioară. În: DJAMGOZ MBA, Archer SN, Varallerga s, editori. Neurobiologie și aspecte clinice ale retinei exterioare. Londra: Chapman & Sala; 1995. p. 395-416.
Robinson SR, Hampson ECGM, Munro MN, Vaney DI. Cuplarea unidirecțională a joncțiunilor gap între neuroglia. Proc. Austr. Neuroști. Soc. 1993;3:167.
Schnitzer J. astrocite în retina mamiferelor. Prog. Ret. Res. 1988; 7: 209-232.
Piatra J, Makarov F, Hollander H. inserarea glială a colinei soma și axon a celulelor ganglionare retiniene. Vis Neurosci. 12:273–279.
Piatra J, Dreher Z. relația dintre astrocite, celule ganglionare și vasculatura retinei. J Comp Neurol. 1987;255:35–49.
Trivino a, Ramirez JM, Salazar JJ, Ramirez ai, Garcia-Sanchez J. studiul imunohistochimic al astrogliei capului nervului optic uman. Viziune Res. 1996; 36: 2015-2028.
Turner DL, Cepko CL. Un progenitor comun pentru neuroni și glia persistă în retina șobolanului târziu în dezvoltare. Natura. 1987;328:131–136.