gliaceller i näthinnan av Helga Kolb
Helga Kolb
tre grundläggande typer av gliaceller finns i den mänskliga näthinnan, Muller-celler, astroglia och mikroglia. Alla beskrevs för näthinnan av Cajal för mer än hundra år sedan (1892).
1. Muller-celler.
Muller-celler är den huvudsakliga glialcellen i näthinnan. De bildar arkitektoniska stödstrukturer som sträcker sig radiellt över näthinnans tjocklek och är gränserna för näthinnan vid respektive yttre och inre begränsande membran. En fullständig förståelse av formen på en Muller-cell ses bäst efter Golgi-färgning som ursprungligen visades av Cajal (1892) nedan.
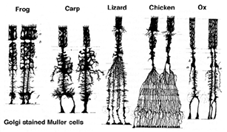
Fig. 1. Golgi-färgade Muller-celler
Muller-cellkroppar sitter i det inre kärnskiktet och projicerar oregelbundet tjocka och tunna processer i båda riktningarna till det yttre begränsande membranet och till det inre begränsande membranet. Muller-cellprocesser insinuerar sig mellan cellkroppar i neuronerna i kärnskikten och kuvertgrupper av neurala processer i de plexiforma skikten (Fig. 1). I själva verket är retinala neurala processer endast tillåtna direktkontakt, utan att omsluta Muller-cellprocesser, vid deras synapser.
en enda stamcell ger upphov till både Muller-celler och retinala neuroner(Turner och Cepko, 1987) men tydligen i två faser. De tidigaste fasneuronerna födda vid den apikala marginalen hos neuroepitelet intill pigmentepitelet producerar primära neuroner som består av konceller, horisontella celler och ganglionceller (Fig. 2, höger). Den andra fasen av celler som också är födda vid apikala marginaler producerar Muller-celler och stavfotoreceptorer, bipolära celler och amakrinceller (Reichenbach och Robinson, 1995) (Fig. 2, vänster). Alla utvecklande neuroner och Muller-cellerna måste migrera inåt till sin slutliga position och man tror att Muller-cellprocesserna och stammarna styr mycket av neuronmigrationerna och styr neurit-differentieringarna.
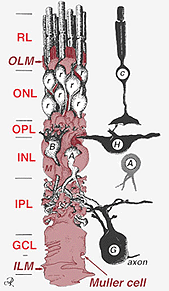
Fig. 2. 3-D schematisk ritning av förhållandet mellan Muller cell och andra retinala neuroner
korsningarna som bildar det yttre begränsande membranet är mellan Muller-celler och andra Muller-celler och fotoreceptorceller som robusta desmosomer eller zonula adherens. I vissa arter är gapkorsningar (specialiserade membranföreningar och kanaler som tillåter passage av små molekyler och joner) eller täta korsningar en del av dessa Muller-cellkorsningar (Miller och Dowling, 1970) men inte så hos däggdjursarter där ingen färgkoppling någonsin har observerats (Robinson et al., 1993; Reichenbach och Robinson, 1995). Ytan på Muller-cellen som vetter mot pigmentepitelet och subretinalutrymmet expanderas av många utsprång av Muller-cellmembranet som kallas apikal villi. Det inre begränsande membranet å andra sidan bildas av Muller-cellens koniska ändfötter men inga specialiserade korsningar ses här. Muller-celler bildar också ändfötter på de stora retinala blodkärlen vid näthinnans inre yta. Ytan på Muller-cellmembranet som vetter mot glaskroppen är täckt med ett mukopolysackaridmaterial och bildar således ett riktigt källarmembran.
Muller-celler innehåller glykogen, mitokondrier och mellanliggande filament som är immunoreativa för vimentin och till viss del till glial fibrillär syraprotein (GFAP). De senare filamenten är normalt i den inre halvan av retinala Muller-cellerna och deras ändfötter (Fig. 3, vänster), men efter trauma på näthinnan, såsom retinalavlossning, uppregleras både vimentin och GFAP massivt och finns i hela cellen (Fig. 3, höger) (Guerin et al., 1990; Fisher och Lewis, 1995).
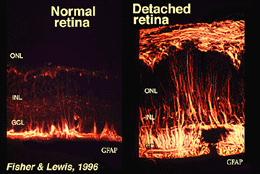
Fig. 3. GFAP-immunoreaktivitet i Muller-celler
Muller-celler har en rad funktioner som alla är avgörande för retinala neurons hälsa. Muller-celler fungerar i ett symbiotiskt förhållande med neuronerna (för en utmärkt recension se Reichenbach och Robinson, 1995). Således inkluderar Muller-cellfunktioner:
- levererar slutprodukter av anaerob metabolism (nedbrytning av glykogen) till bränsle aerob metabolism i nervcellerna.
- de Moppar upp neurala avfallsprodukter som koldioxid och ammoniak och återvinner förbrukade aminosyrasändare.
- de skyddar neuroner från exponering för överskott av neurotransmittorer som glutamat med hjälp av välutvecklade upptagningsmekanismer för att återvinna denna sändare. De kännetecknas särskilt av närvaron av höga koncentrationer av glutaminsyntas.
- de kan vara involverade i både fagocytos av neuronala skräp och frisättning av neuroaktiva ämnen som GABA, taurin och dopamin.
- de tros syntetisera retinsyra från retinol (retinsyra är känd för att vara viktig i utvecklingen av ögat och nervsystemet) (Edwards, 1994)
- de kontrollerar homeostas och skyddar neuroner från skadliga förändringar i deras Joniska miljö genom att ta upp extracellulär K+ och omfördela den.
- de bidrar till genereringen av elektroretinogrammet (ERG) b-våg (Miller och Dowling, 1970; Newman och Odette, 1984), den långsamma P3-komponenten i ERG (Karwoski och Proenza, 1977) och scotopic threshold response (STR) (Frishman och Steinberg, 1989). De gör det genom reglering av k + distribution över retinal glaskroppen gränsen, över hela näthinnan och lokalt i det inre plexiform skiktet av näthinnan (Fig. 4, från Reichenbach och Robinson, 1995, anpassad från Newman, 1989).

Fig. 4. Reglering av K + av Muller-celler i Muller-celler
2. Astrocyter.
astrocyter är inte gliaceller i retinalneuroepitelet utan kommer in i den utvecklande näthinnan från hjärnan längs den utvecklande optiska nerven (Stone and Dreher, 1987; Chan-Ling 1994). De har en karaktäristisk morfologi av en utplattad cellkropp och en fibrös serie strålningsprocesser. Mellanliggande filament fyller sina processer och därmed fläckar de dramatiskt med antikroppar mot GFAP (Schnitzer, 1988). Astrocytcellkroppar och processer är nästan helt begränsade till nervfiberskiktet i näthinnan. Deras morfologi förändras från periferin till det optiska nervhuvudet: från en symmetrisk stellatform i perifer näthinna (Fig. 5a och b) (Schitzer, 1988) till extremt långsträckt nära optisk nerv (Fig. 6 och 7).
 Fig. 5a. astrocyter i perifer näthinna. Schnitzer, 1988 |
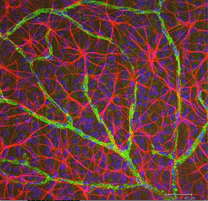 Fig. 5b. Astrocyter (röda) i råtthinnan bland ganglionceller (blå) och associerade med blodkärl (grön). Med tillstånd av Laura Fernandez-Sanchez och Nicolas Cuenca |
i immunocytokemisk färgning (Fig. 5b) och i HRP intracellulära injektioner (Fig. 7) färgade astrocyter uppvisar tydligt processer inriktade längs ganglioncellsaxonerna som går igenom nervfiberskiktet. Vid distribution når astrocyter sin topp på det optiska nervhuvudet och har en ganska jämn minskning av densiteten i utstrålande ringar från nervhuvudet. De är inte närvarande i avaskulär fovea eller ora serrata.
 Fig. 6. Astrocyter i centrala näthinnan. Schnitzer, 1988 |
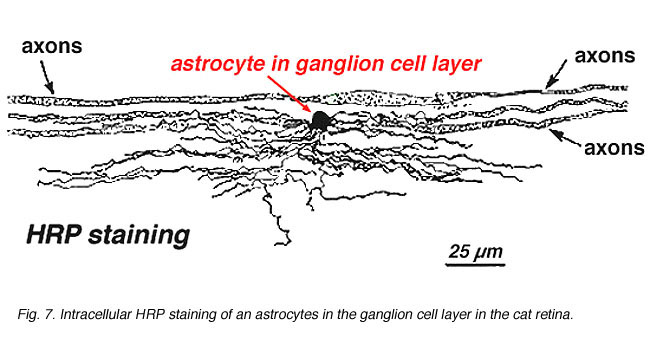 Fig. 7. Astrocyter i katt näthinnan färgas genom intracellulär injektion av HRP. Med tillstånd av Ralph Nelson. |
tjocka och tunna astrocyter har särskiljats på grundval av färgning med antikroppar mot GFAP (Trevino et al., 1996). Således är astrocyter anordnade över ytan av ganglioncellsaxonbuntarna när de går in i det optiska nervhuvudet och bildar ett rör genom vilket axonerna löper (Fig. 8). Gap-korsningar och zonula-adherens-korsningar har beskrivits mellan astrocytiska processer i katt näthinnan (h Obbllander et al., 1991).
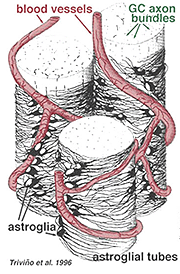
Fig. 8. 3-D-block av astrocyter anordnade över ytan av ganglioncell axonbuntar
blodkärlen som körs i och bland ganglioncellbuntarna täcks också av båda processerna och till och med en tillfällig cellkropp av en astrocyt. Funktionen hos astrocyter som omsluter ganglioncellsaxoner och förhållandet till blodkärlen i nervfiberskiktet antyder att de är axonala och vaskulära glialmantlar och en del av en blod-hjärnbarriär. I likhet med Muller-celler är de kända för att innehålla riklig glykogen och de kan bilda en näringstjänst för att ge glukos till neuronerna. Dessutom tjänar de förmodligen en roll i jonisk homeostas vid reglering av extracellulära kaliumnivåer och metabolism av neurotransmittorer som GABA.
3. Mikrogliaceller.
den tredje glialcellstypen är förmodligen av mesodermalt ursprung och är således strängt taget inte neuroglial liksom astrocyterna och Muller-cellerna. De kommer in i näthinnan som sammanfaller med de mesenkymala prekursorerna av retinala blodkärl under utveckling (Chan-Ling, 1994). Mikroglialceller är allestädes närvarande i den mänskliga näthinnan som finns i varje lager av näthinnan.
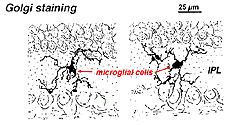
i Golgi-färgad näthinna ser de ut som konstiga, multipolära former med små cellkroppar och oregelbundna korta processer. I Golgi-preparat har de faktiskt ibland misstagits för nervceller, särskilt när de ligger i ett kärnskikt med en enda orientering av sina processer i det plexiforma skiktet.
Fig. 9. Golgifärgning av mikroglialceller
mikroglialceller kan vara av två typer. En form tros komma in i näthinnan i tidiga utvecklingsstadier från det optiska nervmesenkymet och ligga vilande i näthinnskikten under mycket av näthinnans liv. Den andra formen av mikroglia verkar vara blodburna celler, möjliga härrörande från kärlpericyter (Boycott och Hopkins, 1981; Gallego, 1986). Båda typerna kan stimuleras till en makrofagisk funktion efter trauma mot näthinnan, och sedan engagerar de sig i fagocytos av degenererande retinala neuroner.

Fig. 10a. Lektinfärgad mikroglialcell från Chan-Lin, 1994.
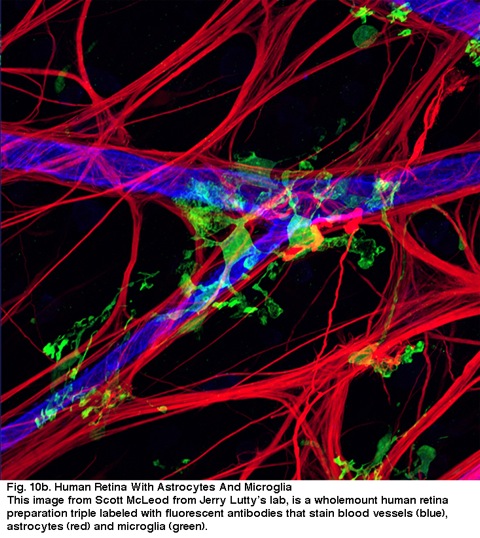
Fig. 10b. mänsklig näthinna med astrocyter och mikroglia. Denna bild från Scott McLeod från Jerry Lutty ’ S lab, är en helhetlig Human retina-beredning trippel laneled med fluorescerande antikroppar som fläckar blodkärl (blå), astrocyter (röd) och mikroglia (grön).
4. Hänvisningar.
bojkott BB, Hopkins JM. Mikroglia i näthinnan hos apa och andra däggdjur; dess skillnad från andra typer av glia och horisontella celler. Neurovetenskap.1981;6:679–688.
Cajal SR. In: Thorpe SA, Glickstein M, översättare. 1892. Strukturen av näthinnan. Springfield (IL): Thomas; 1972.
Chan-Ling T. Gliala, neuronala och vaskulära interaktioner i däggdjurs näthinnan. Prog. Ret. Öga Res. 1994; 13: 357-389.
Edwards RB. Biosyntes av retinsyra av M Ellereller gliaceller: en modell för centrala nervsystemet? Prog. Ret. Öga Res. 1994; 13: 231-242.
Fisher SK, Lewis GP. Fotoreceptorer och bortom: cellulära och molekylära effekter av retinalavlossning.2: A Great Basin Visual Science Symposium, II, University of Utah Press. 1995
Frishman LJ, Steinberg RH. Ljus framkallade ökningar i o i proximala delen av den mörka anpassade katt näthinnan. J Neurofysiol. 1989;61:1233–1243.
Gallego A. jämförande studier på horisontella celler och en anteckning på mikroglialceller. Prog. Ret. Res. 1986; 5: 165-206.
Guerin CJ, Anderson DH, Fisher SK. Förändringar i intermediär filamentimmunmärkning sker som svar på retinalavlossning och återmontering i primater.Investera. Oftal. Vis. Sci. 1990;31:1474–1482.
Karwoski CJ, Proenza LM. Förhållandet mellan Muller-cellsvar, en lokal transretinal potential och kaliumflöde. J Neurofysiol. 1977;40:244–259.
Miller RF, Dowling JE. Intracellulära svar av Muller (glial) celler i mudpuppy retina: deras förhållande till B-våg av elektroretinogrammet. J Neurofysiol.1970;33:323–341.
Newman EA, Odette LL. Modell av elektroretinogram B-våggenerering: ett test av k+ – hypotesen. J Neurofysiol. 1984;51:164–182.
Newman EA. Elektrofysiologi av retinala glialceller. Prog. Ret. Res. 1989; 8: 153-172.
Reichenbach A, Robinson SR. Inblandning av M-celler i den yttre näthinnan. I: Djamgoz MBA, Archer SN, Valler S, redaktörer. Neurobiologi och kliniska aspekter av den yttre näthinnan. London: Chapman & Hall; 1995. s. 395-416.
Robinson SR, Hampson ECGM, Munro MN, Vaney DI. Enkelriktad koppling av gapkorsningar mellan neuroglia. Proc. Austr. Neurovetenskap. Soc. 1993;3:167.
Schnitzer J. astrocyter i näthinnan hos däggdjur. Prog. Ret. Res. 1988; 7: 209-232.
Sten J, Makarov F, Hollander H. glial ensheathment av Soma och axon hillock av retinala ganglionceller. Vis Neurosci. 12:273–279.
Sten J, Dreher Z. förhållandet mellan astrocyter, ganglionceller och kärl i näthinnan. J Comp Neurol. 1987;255:35–49.
Trivino A, Ramirez JM, Salazar JJ, Ramirez AI, Garcia-Sanchez J. immunhistokemisk studie av mänsklig optisk nervhuvud astroglia. Vision Res. 1996; 36: 2015-2028.
Turner DL, Cepko CL. En vanlig stamfader för neuroner och glia kvarstår i råtta näthinnan sent i utvecklingen. Natur. 1987;328:131–136.