frontiere în chimie
- Introducere
- materiale și metode
- materiale
- culturi celulare
- Setări imagistice SPR bazate pe prismă
- pregătirea cipului senzorului
- sistem de curgere
- calibrarea sensibilității
- rezultate și discuții
- pierderea inițială a densității de masă în fixarea celulară prin PFA
- răspunsul presiunii osmotice în fixarea celulară de către PFA
- pierderea substanțială a densității de masă în permeabilizarea celulară de către Triton
- concluzie
- contribuțiile autorului
- Declarație privind conflictul de interese
- mulțumiri
Introducere
imunofluorescența este o tehnică puternică de vizualizare a distribuției biomoleculelor specifice în probele biologice, cum ar fi celulele și țesuturile (Joshi și Yu, 2017). În testele imunofluorescente tipice pe bază de celule, celulele aderente au fost incubate cu anticorp fluorescent pentru a permite recunoașterea și legarea specifică între anticorp și molecula țintă din celule. După clătirea temeinică, proba a fost plasată sub microscop fluorescent pentru a obține o imagine fluorescentă, din care distribuția moleculelor țintă a fost raportată de etichetele fluorescente. Pentru a facilita accesibilitatea anticorpilor la moleculele țintă și pentru a inhiba activitatea celulară inerentă, probele au fost adesea fixate și permeabilizate înainte de procedurile de colorare. Acestea au fost deosebit de necesare atunci când moleculele țintă au fost localizate în citoplasmă. Printre multe tipuri de reactivi, paraformaldehida (PFA) și Triton X-100 sunt probabil cele mai utilizate pentru fixare și, respectiv, permeabilizare. Depolimerizarea PFA a produs molecule de formaldehidă pentru a crea legături chimice covalente între proteinele din probă. Mecanismul acțiunii sa bazat pe activarea unuia dintre reziduurile de aminoacizi lizină. PFA a dizolvat, de asemenea, unele lipide în membranele celulare care au deteriorat ușor integritatea membranei celulare (Fox și colab., 1985; Thavarajah și colab., 2012; Kiernan, 2018). Triton X-100, ca detergent neionic eficient, ar putea dizolva lipidele din membranele celulare, astfel încât membrana celulară a devenit mai permeabilă la anticorpul fluorescent. Etapa de permeabilizare a îndepărtat mai multe lipide ale membranei celulare datorită grupărilor sale de cap hidrofile neîncărcate, care constau din fragmente de polioxietilenă pentru a permite moleculelor mari, cum ar fi anticorpii, să pătrundă în celulă (Jamur și Oliver, 2010; Koley și Bard, 2010). O ilustrare schematică a procesului de fixare și permeabilizare a celulelor este prezentată în figura 1D. În ciuda faptului că fixarea și permeabilizarea au devenit proceduri de rutină în imunofluorescență și imunohistochimie, s-au făcut eforturi destul de rare pentru a clarifica cantitativ influențele lor asupra integrității membranei și densității masei celulare la nivel unicelular.
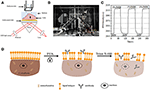
Figura 1. Prism bazate pe SPR Imaging setari. (A) ilustrarea schematică a ansamblului experimental. (B) o fotografie a aparatului. (C) curba de calibrare a sensibilității. (D) ilustrarea schematică a fixării și permeabilizării celulelor.
rezonanța plasmonului de suprafață (SPR) a fost o tehnică populară și puternică pentru a determina cinetica de legare între o pereche de molecule de la inventarea sa în anii 1980 (liedberg și colab., 1983; Cullen și colab., 1987). Acest lucru este atribuit avantajelor sale remarcabile, inclusiv măsurarea cinetică cantitativă în timp real cu rezoluții temporale ridicate, compatibilitatea cu dispozitivele extinse și, cel mai important, caracteristica sa intrinsecă a etichetei fără etichete (Rothenhausler și Knoll, 1988; Phillips și Cheng, 2007; Homola, 2008; Abadian și colab., 2014; m., 2014; Yanase și colab., 2014; Su și Wang, 2018). Aplicațiile timpurii s-au concentrat în principal pe caracterizarea și cuantificarea interacțiunilor biomoleculare prin imobilizarea moleculelor purificate pe substraturile de detectare a SPR (lamele de sticlă acoperite cu aur). Astfel de studii ex situ nu numai că au necesitat proceduri de purificare intensivă a forței de muncă, dar au condus și la rezultate care ar putea să nu reflecte interacțiunea naturală la ființele vii (Phillips și Cheng, 2007; Homola, 2008). Condus atât de progresul tehnic în diferite sisteme de imagistică SPR (SPRi), cât și de motivația științifică în imagistica și analiza cu o singură celulă, atât prism, cât și tehnici SPRi bazate pe obiective au fost folosite pentru a imagina distribuția în masă a celulelor vii unice fără a fi nevoie de etichetare (Wang și colab., 2010, 2012a; Yanase și colab., 2010; Yang și colab., 2015; Zhang și colab., 2015). Fiecare are propriile sale merite. Acesta din urmă are o rezoluție spațială mai mare, iar primul are o sensibilitate mai bună. Prin monitorizarea imaginilor SPR expirate în timp ale celulelor vii unice în timpul diferitelor tipuri de stimulări fiziologice și biologice, informații spațiale și dinamice importante privind interacțiunile celulă-substrat (Giebel și colab., 1999), migrațiile celulare (Smith și colab., 2004), răspunsuri osmotice (Wang și colab., 2012a), cinetica de legare a ligandului-receptor (Wang și colab., 2012b, 2014), căi de semnalizare (Deng și colab., 2016), dinamica activării proteinelor (Peng și colab., 2018) și răspunsurile la medicamentele cu celule canceroase vii (Wang și colab., 2018) au fost obținute. Pentru cazuri, studiile existente au demonstrat în mod clar că tehnicile SPRi au fost capabile să cartografieze densitatea de masă, precum și integritatea membranei la nivel de celulă unică (Yanase și colab., 2010; Wang și colab., 2012b; Yang și colab., 2015). Contrastul imaginii SPRi a venit din distribuția subcelulară a indicelui de refracție, care a fost determinată aproximativ de densitatea de masă locală. Legarea anticorpilor specifici pe membrana celulară care a exprimat antigenii corespunzători sau stimularea celulelor vii cu anumite substanțe chimice s-a dovedit a modifica densitatea de masă a celulelor unice într-o manieră eterogenă și dinamică (Wang și colab., 2012a, b). S-a constatat că expunerea celulelor vii unice la soluții hipertonice induce contracția celulelor, indicând integritatea excelentă a membranelor celulare (Wang și colab., 2012a). Cu toate acestea, modul în care tratamentele de fixare și permeabilizare ar afecta imaginile SPR ale celulelor unice rămâne neclar.
în lucrarea de față, am folosit un sistem SPRi construit acasă pentru a înregistra continuu imaginile SPR expirate în timp a zeci de celule vii unice atunci când celulele au fost tratate succesiv cu 4% soluție PFA și 1% soluție Triton X-100. Densitatea de masă a celulelor unice a fost determinată de semnalul SPRi mediu. Integritatea membranei a fost evaluată prin expunerea celulelor la soluție hipertonică. S-a constatat că, pe lângă densitatea de masă ușor redusă cu <10%, tratamentul cu PFA a distrus semnificativ integritatea celulară, așa cum este indicat de pierderea răspunsului osmotic la expunerea moleculelor de zahăr. Tratamentul ulterior cu Triton X-100 a redus semnificativ densitatea de masă cu încă 20%, sugerând distrugerea severă a integrității membranei.
materiale și metode
materiale
soluție salină tamponată cu fosfat Dulbecco (PBS, Gibco), zaharoză (Sinopharm Chemical Reactiv Co., Ltd), Triton X-100 (Aladdin), Paraformaldehidă (Shanghai Lingfeng Chemical Reactiv Co., Ltd). Toți reactivii au fost dizolvați în PBS.
culturi celulare
celulele BT-474 au fost cultivate la 37 CTC cu 5% CO2 și 70% umiditate relativă în mediul vulturului modificat Dubelco (DMEM, Invitrogen) cu 10% ser fetal bovin (FBS, Invitrogen), 100 Unități/mL penicilină și 100 hectolitri / mL streptomicină (Invitrogen). Celulele au fost transmise când au fost 70-80% confluente prin tratarea cu soluție de tripsină 0,25% (Gibco).
Setări imagistice SPR bazate pe prismă
o diagramă schematică și o fotografie a sistemului SPRi sunt prezentate în figurile 1a,B. aparatul SPRi este compus în principal din trei părți: sursă de lumină, componente optice și cameră foto.
• sursă de lumină: o diodă emițătoare de lumină de 670 nm (L7868-01, Hamamatsu, Japonia) și o priză de montare cu temperatură controlată (LDM21, Thorlabs, Newton, SUA).
• componente optice (Edmund Optics, SUA): o prismă triunghiulară SF-11 cu cip senzor pornit (nr. 1 sticlă BK7 de la Fisherbrand), o lentilă convergentă, un polarizator și o lentilă reglabilă.
• cameră: cameră cu dispozitiv cuplat la Încărcare (Pike F032B, Allied Vision Technology, Newburyport, SUA).
lumina polarizată p lovește direct partea inferioară a diapozitivului de sticlă acoperit cu aur prin prisma și camera captează fasciculul reflectat pentru a genera imagini SPR la o rată de 0,828 cadru pe secundă (fps). Atunci când alegeți o mărire de 4x într-o lentilă reglabilă, câmpul vizual a fost de ~2,2 mm2.
pregătirea cipului senzorului
fiecare cip a fost spălat cu 75% etanol și apă deionizată (DIW), urmată de expunerea la UV timp de 30 de minute pentru a îndepărta contaminarea suprafeței și a le steriliza înainte de fiecare experiment. O cameră de siliciu Flexi-Perm (Greiner Bio-One) a fost plasată deasupra cipului de aur pentru a servi ca o cultură celulară bine. Pentru a realiza modificarea suprafeței înainte de însămânțarea celulară, în cameră s-a adăugat o soluție de colagen de 100 hectog/mL 150 centicl și s-a păstrat într-un incubator timp de 2 ore. cipul a fost apoi clătit cu apă deionizată de două ori și DMEM de două ori înainte de însămânțarea celulară. Pentru a face celulele o formă bună, cipul a fost incubat în mediul de creștere timp de 36 h. mediul de creștere a fost apoi înlocuit cu soluție tampon PBS timp de 10 minute pentru ca celulele să obțină un echilibru.
sistem de curgere
un sistem de perfuzie medicament multicanal acționat gravitațional a fost aplicat la injectarea și comutarea soluției. Debitul este de 300 unqql / min, iar viteza de comutare a soluției în jurul celulelor poate ajunge la 1-2 s.
calibrarea sensibilității
a fost pregătit un cip senzor așa cum este descris în partea precedentă fără modificarea suprafeței și însămânțarea celulelor. Mai întâi, DIW curgea peste cip timp de aproximativ 60 s. apoi, a fost înlocuit cu soluție de etanol 1% (v/v) pentru încă 60 s. ca una dintre cele mai frecvent utilizate soluții de calibrare, se știe că 1% etanol crește unghiul SPR al DIW cu 60 mDeg. Rezultatele experimentale au arătat că intensitatea SPR a crescut în consecință cu 96 U. I. (figura 1C), corespunzând unei sensibilități de 1,6 UQTI.U. / mDeg.
rezultate și discuții
membrana celulară (cunoscută și sub numele de membrană citoplasmatică), constând dintr-o bistrat lipidic cu proteine încorporate, este o membrană semipermeabilă care cuprinde citoplasma unei celule. Etapele de fixare și permeabilizare a celulelor și a probelor de țesut, care ar putea modifica permeabilitatea membranei celulare, sunt proceduri cruciale care ar putea determina succesele testelor imunofluorescente sau imunohistochimice. În plus, calitatea anticorpilor și procedura de imunoreacție sunt alți factori determinanți cheie pentru aceste tipuri de experimente. SPR are caracteristica că atât unghiul rezonant, cât și indicele de refracție (RI) în apropierea suprafeței de detectare sunt foarte sensibile la densitatea de masă a stratului de suprafață din interfața mediu-metal. Prin urmare, variațiile densității de masă a celulei care au loc pe sau în apropierea filmului metalic (~200 nm) ar putea fi înregistrate sincron prin măsurarea intensității luminii reflectate. Prin utilizarea tehnicii de imagistică prin rezonanță a plasmonului de suprafață, am explorat distribuția dinamică a densității masei celulare cu rezoluții spațiale și temporale ridicate și sensibilitate fiabilă atât în procesele de fixare, cât și în cele de permeabilizare în acest studiu.
pierderea inițială a densității de masă în fixarea celulară prin PFA
prima etapă de pregătire a probelor biologice pentru analiza imunofluorescentă sau imunohistochimică este de obicei fixarea. Și cele mai frecvent utilizate fixative pentru aceste tipuri de teste sunt fixative de reticulare precum PFA care funcționează prin generarea de legături chimice covalente între proteinele din celule sau țesuturi. Prin urmare, am investigat influența tratamentului PFA asupra densității masei celulare. Celulele vii BT-474 au fost cultivate pe lamele acoperite cu folie de aur cu o grosime de 50 nm. O imagine SPR reprezentativă care conține celule unice este prezentată în figura 2a. semnalele SPR ale fiecărei celule pot fi obținute prin alegerea unei regiuni de interes din imaginea SPR care să corespundă morfologiei celulei investigate. Într-un experiment tipic, când soluția curgea peste celule, s-au obținut senzori SPR ai celulelor analizând fiecare celulă așa cum se arată în figura 2b (curba neagră). În precedentele 242 s, tamponul PBS fără PFA curgea peste celule, iar linia de bază a fost determinată de stabilitatea sistemului (sursa de lumină și stabilitatea mecanică) și de semnalele inerente de micromotie generate de celulele vii. Soluția PFA a fost apoi introdusă la a 242-a secundă, ceea ce a crescut imediat semnalul SPR din cauza indicelui de refracție relativ mare de soluție PFA 4% (Efect în vrac). Soluția PFA a funcționat încă 17 minute pentru a obține o fixare suficientă. La a 1237-a secundă, soluția PFA a fost comutată înapoi la soluția PBS pură. În plus față de efectul redus în vrac, s-a constatat o scădere a intensității SPR a celulelor unice atunci când s-au comparat semnalele înainte (0-242) și după (1237-1545) tratamentele PFA și procentul de pierdere de masă a fost obținut prin împărțirea valorii de scădere la valoarea inițială. Sensongramele SPR ale fundalului (o regiune adiacentă fără adeziune celulară, curbă albastră) și o singură celulă tratată cu PBS în timpul 1.500 s (curbă gri) sunt de asemenea prezentate în figura 2b. analiza statistică pe 30 de celule relevă o pierdere medie a densității de masă de 10% 5%, așa cum se arată în figura 2c. De asemenea, s-a constatat că, după tratamentul cu PFA, nivelul de fluctuație de bază în semnalul SPR a fost redus semnificativ la jumătate sau chiar la două treimi, sugerând pierderea micromotiilor și fixarea celulelor tratate așa cum se arată în figura 2D. astfel de semnale de micromotie au fost generate de o grămadă de activități celulare dinamice din diferite componente celulare, inclusiv schelet, membrane și organite. Fixarea a încheiat astfel de activități biologice și, prin urmare, a eliminat semnalele de micromotie, însoțind cu o fluctuație de intensitate semnificativ redusă în sensogramele SPR ale celulelor unice.
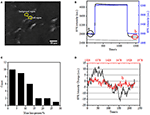
Figura 2. Procesul de fixare a celulelor. (A) Imaginea SPRi a celulelor BT-474. (B) semnalele SPR ale celulei unice (curba neagră) și ale fundalului (curba albastră) supuse unui proces de comutare a soluției: tampon PBS la 4% tampon PFA la PBS. Semnal SPR al unei singure celule tratate cu PBS în timpul 1.500 s (curba gri). (C) distribuția pierderilor de masă a 30 de celule unice. (D) nivelul de zgomot al celulei înainte de (a) și după (B) tratamentul PFA.
răspunsul presiunii osmotice în fixarea celulară de către PFA
structura integrată a membranei celulare, o membrană semipermeabilă, reglează homeostazia unei celule pentru a atinge echilibrul în presiunea osmotică fiziologică. S-a constatat, de asemenea, că tratamentul cu PFA are capacitatea de a distruge integritatea membranei așa cum se arată în Figura 3. Când tamponul din jurul celulei țintă s-a schimbat în soluție de zaharoză de 25 mM în <1 s printr-un sistem de perfuzie medicamentoasă, a fost creat un mediu de cultură celulară hipertonică. Pentru celulele vii, expunerea la soluția hipertonică a declanșat o serie de răspunsuri fiziologice pentru a echilibra presiunea osmotică intracelulară și extracelulară, care a fost investigată în detaliu în lucrarea noastră anterioară (Wang și colab., 2012a). În acest studiu, un sensorgram tipic SPR al celulelor BT-474 fiziologice unice este furnizat în figura 3a. la expunerea la soluția de zaharoză hipertonică, celulele tratate au suferit mai întâi o contracție presantă, ducând la o creștere treptată a intensității SPR până la atingerea unui platou. Acesta a sugerat creșterea densității de masă ca urmare a contracției celulare și a componentelor subcelulare care se adună spre fundul capacului culturii de aur. Creșterea treptată a intensității SPR prin stimulare hipertonică este un fel de reglare fiziologică a presiunii osmotice celulare, indicând astfel integritatea fiziologică a membranei celulare. Când integritatea membranei a fost distrusă, s-a constatat că introducerea soluției de zaharoză hipertonică crește imediat intensitatea SPR într-un timp mult mai scurt, așa cum se arată în figura 3b. Aceasta se datorează faptului că moleculele de zahăr au pătruns liber în celule fără rezistență din membrana semipermeabilă și au crescut imediat indicele de refracție local (efect în vrac). Este clar că, în timp ce creșterea lentă a intensității SPR reflecta reglarea osmotică fiziologică (figura 3a), saltul rapid a demonstrat că membrana celulară a fost deteriorată pentru a permite intrarea liberă a moleculelor mici de zahăr (figura 3b).

Figura 3. Răspunsurile la presiunea osmotică a celulelor. (A) Răspunsul unei celule vii la zaharoză de 25 mM. (B) răspunsul aceleiași celule fixe (10 min) la 25 mM zaharoză.
pierderea substanțială a densității de masă în permeabilizarea celulară de către Triton
în etapa de fixare anterioară, o densitate inițială a masei celulare a scăzut doar cu ~10% (figura 2c). Este rezonabil să speculăm că fixarea PFA a distrus doar ușor integritatea membranei și a crescut permeabilitatea moleculelor mici. Majoritatea componentelor intracelulare trebuie încă adăpostite în membrana citoplasmatică. Ulterior am studiat influența unui detergent tipic de permeabilizare, 1% Triton X-100, asupra celulelor care au fost fixate cu PFA timp de 17 minute anterior. Un sensorgram SPR reprezentativ al celulelor unice în timpul expunerii la 1% Triton X-100 este prezentat în figura 4a. introducerea Triton X-100 a crescut intensitatea SPR urmată de o scădere treptată. Prima creștere este o consecință a creșterii indicelui de refracție al soluției în vrac, iar scăderea urmată a indicat pierderea treptată a densității de masă datorată permeabilizării celulare. Densitatea de masă redusă a fost mai abundentă atunci când se compară modificarea intensității SPR înainte (0-300 s) și după introducerea detergentului (1.300–1.800 s). Analiza pe 30 de celule unice a demonstrat că tratamentul cu Triton X-100 a creat o scădere substanțială a densității masice cu încă 20 de centi 5% după fixarea PFA, așa cum se arată în figura 4D și în tabelul 1. Este clar că detergentul Triton X-100 a eliminat mult mai multe lipide ale membranei celulare și a distrus grav integritatea membranei în această etapă de permeabilizare. Acest lucru a dus la eliberarea mai multor componente intracelulare, în special macromolecule, din celule. Și, de asemenea, acest lucru permite moleculelor mari, cum ar fi anticorpii, să intre în celulă în următoarea etapă de imunoreacție. Deși intensitatea SPR a levigării moleculelor mici a fost detectată după fixarea PFA, permeabilizarea prin utilizarea detergentului Triton X-100 a solubilizat în mod clar lipidele și a făcut celulele mult mai permeabile la mișcările macromoleculelor în și în afara corpului celular.
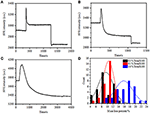
Figura 4. Curbele pierderii de masă ale celulei tratate cu soluții de concentrație diferite (a) 1,0%, (B) 0,5%, (C) 0,1%. (D) distribuția pierderilor de masă a 30 de celule unice în condiții diferite.

Tabelul 1. Un rezumat al variațiilor în pierderea densității de masă a celulelor statistice în diferite condiții.
s-a constatat că pierderea densității de masă este un proces destul de rapid, care a avut loc în câteva minute în stadiul de permeabilizare (figura 4A). Scăderea densității de masă a urmat aproximativ o descompunere monotonă într-o perioadă de timp de 126 s. sensorgramele tipice ale celulelor unice sub 0,5 și 0,1% Triton X-100 sunt afișate în figurile 4b,respectiv C. Procesul de permeabilizare a devenit mult mai lent la reducerea concentrației de detergent. A fost nevoie de 126, 510, 2.600 s pentru cele trei tipuri de celule tratate pentru a termina pierderea de masă.
concluzie
în rezumat, redistribuirea dinamică a densității masei celulare atât în procesele de fixare, cât și în cele de permeabilizare au fost explorate sensibil prin utilizarea configurației SPRi bazate pe prisme cu rezoluții spațiale și temporale ridicate. PFA fixativ ar putea crește inițial permeabilitatea membranei citoplasmatice într-o oarecare măsură, deoarece capacitatea sa de fixare a proteinelor celulare atât în proteine membranare, cât și intra-celulare. A fost susținută de (1) scăderea densității de masă cu ~10% după un tratament de fixare de 17 minute în soluție PFA 4% și (2) răspunsul celular eliminat la soluția hipertonică, care a demonstrat distrugerea integrității celulare la molecule mici precum zaharurile. Detergentul Triton X-100 este superior PFA fixativ în solubilizarea lipidelor și, prin urmare, crește permeabilitatea membranei celulare, însoțind pierderea mai substanțială a densității de masă datorată eliberării nu numai a moleculelor mici, ci și a moleculelor mari și, eventual, a unor organite, cum ar fi veziculele. În plus, rata pierderii de masă este corelată pozitiv cu concentrația Triton X-100. Aceste rezultate au oferit înțelegeri cantitative și dinamice asupra influenței fixării și permeabilizării asupra densității masei celulare și a integrității membranei, cu implicații pentru optimizarea condițiilor pentru experimentele biologice cu o singură celulă, cum ar fi testele imunofluorescente și imunohistochimice.
contribuțiile autorului
RC, Y-WS și WW au proiectat experimentele. RC, FZ, ML și XW au efectuat experimentele. RC, Y-WS și WW au scris lucrarea. RC, FZ, Y-WS și WW au discutat rezultatele și au analizat datele.
Declarație privind conflictul de interese
autorii declară că cercetarea a fost realizată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
editorul de manipulare a declarat o afiliere comună, deși nu există altă colaborare, cu autorii WW, RC, ML și XW la momentul revizuirii.
mulțumiri
mulțumim sprijin financiar de la Fundația Națională de științe Naturale din China (Grant Nr 21522503).
Abadian, P. N., Kelley, C. P. și Goluch, E. D. (2014). Analiza și detectarea celulară utilizând tehnici de rezonanță a plasmonului de suprafață. Anal. Chem. 86, 2799–2812. doi: 10.1021/ac500135s
PubMed Abstract | CrossRef Full Text/Google Scholar
Cullen, D. C., Brown, R. G. W. și Lowe, C. R. (1987). Detectarea formării imuno-complexe prin rezonanța plasmonului de suprafață pe grătarele de difracție acoperite cu aur. Biosenzori 3, 211-225. doi: 10.1016 / 0265-928X (87)85002-2
PubMed Abstract | CrossRef Full Text | Google Scholar
Deng, S., Yu, X., Liu, R., Chen, W., și Wang, P. (2016). Un dispozitiv microfluidic cu două compartimente pentru detectarea pe termen lung a celulelor vii bazate pe rezonanța plasmonului de suprafață. Biomicrofluidics 10: 044109. doi: 10.1063/1.4960487
PubMed Abstract | CrossRef Full Text/Google Scholar
Fox, C. H., Johnson, F. B., Whiting, J. și Roller, P. P. (1985). Fixarea formaldehidei. J. Histochem. Cytochem. 33, 845–853. doi: 10.1177 / 33.8.3894502
PubMed Abstract | CrossRef Full Text/Google Scholar
Giebel, K. F., Bechinger, C., Herminghaus, S., Riedel, M., Leiderer, P., Weiland, U. și colab. (1999). Imagistica contactelor celulare / substratului celulelor vii cu microscopie de rezonanță a plasmonului de suprafață. Biophys. J. 76, 509-516. doi: 10.1016 / S0006-3495 (99)77219-X
PubMed Abstract | CrossRef Full Text | Google Scholar
Homola, J. (2008). Senzori de rezonanță plasmonică de suprafață pentru detectarea speciilor chimice și biologice. Chem. 108, 462-493. doi: 10.1021 / cr068107d
PubMed Abstract | CrossRef Full Text/Google Scholar
Jamur, M. C. și Oliver, C. (2010). „Permeabilizarea membranelor celulare”, în metodele și protocoalele Imunocitochimice, eds C. Oliver și M. C. Jamur (Totowa, NJ: Humana Press), 63-66. doi: 10.1007 / 978-1-59745-324-0_9
CrossRef Text Complet/Google Scholar
Joshi, S. și Yu, D. (2017). „Capitolul 8-imunofluorescența”, în metodele științifice de bază pentru cercetătorii clinici, eds M. Jalali, F. Y. L. Saldanha și M. Jalali (Boston, MA: Academic Press), 135-150. doi: 10.1016 / B978-0-12-803077-6.00008-4
CrossRef Full Text | Google Scholar
Kiernan, J. A. (2018). Formaldehidă, formalină, paraformaldehidă și glutaraldehidă: ce sunt și ce fac. Micros. Astăzi 8, 8-13. doi: 10.1017 / S1551929500057060
CrossRef Full Text/Google Scholar
Koley, D. și Bard, aj (2010). Efectele concentrației Triton X-100 asupra permeabilității membranei unei singure celule HeLa prin scanarea microscopiei electrochimice (SECM). Proc. Natl. Acad. Sci. SUA 107, 16783-16787. doi: 10.1073 / pnas.1011614107
PubMed Abstract | CrossRef Full Text/Google Scholar
Liedberg, B., Nylander, C., și Lunstr, I. (1983). Rezonanța plasmonului de suprafață pentru detectarea și biosenzarea gazelor. Sens. Actionat. 4, 299–304. doi: 10.1016/0250-6874(83)85036-7
CrossRef Full Text | Google Scholar
m Inktivjard, R., Griesser, H. J. și Thierry, B. (2014). Biosenzare optică pentru studii celulare fără etichete. TrAC Tendințe Anal. Chem. 53, 178–186. doi: 10.1016 / j. trac.2013.08.012
CrossRef Text Complet/Google Scholar
Peng, Z., Lu, J., Zhang, L., Liu, Y., și Li, J. (2018). Imagistica fără etichetă a răspunsului indus de receptorul factorului de creștere epidermal în celule vii unice. Analist 143, 5264-5270. doi: 10.1039 / C8AN01534A
PubMed Abstract | CrossRef Full Text/Google Scholar
Phillips, K. S., și Cheng, Q. (2007). Progrese recente în tehnicile bazate pe rezonanța plasmonului de suprafață pentru bioanaliză. Anal. Bioanal. Chem. 387, 1831–1840. doi: 10.1007 / s00216-006-1052-7
PubMed Abstract | CrossRef Full Text/Google Scholar
Rothenhluxusler, B., și Knoll, W. (1988). Microscopie de suprafață-plasmon. Natură 332, 615-617. doi: 10.1038 / 332615a0
CrossRef Full Text/Google Scholar
Smith, J. T., Tomfohr, J. K., Wells, M. C., Beebe, T. P., Kepler, T. B. și Reichert, W. M. (2004). Măsurarea migrației celulare pe gradienții de fibronectină legați la suprafață. Langmuir 20, 8279-8286. doi: 10.1021/la0489763
PubMed Abstract | CrossRef Full Text/Google Scholar
Su, yw, și Wang, W. (2018). Detectarea rezonanței plasmonului de suprafață: de la biomolecule purificate la celule intacte. Anal. Bioanal. Chem. 410, 3943–3951. doi: 10.1007 / s00216-018-1008-8
PubMed Abstract | CrossRef Full Text/Google Scholar
Thavarajah, R., Mudimbaimannar, V. K., Elizabeth, J., Rao, Marea Britanie și Ranganathan, K. (2012). Elementele de bază chimice și fizice ale fixării de rutină a formaldehidei. J. Maxilofac Oral. Pathol. 16, 400–405. doi: 10.4103/0973-029X.102496
PubMed Abstract | CrossRef Full Text/Google Scholar
Wang, S., Shan, X., Patel, U., Huang, X., Lu, J., Li, J. și colab. (2010). Imagistica, detectarea și măsurarea masei fără etichete a virusurilor unice prin rezonanța plasmonului de suprafață. Proc. Natl. Acad. Sci. SUA 107, 16028-16032. doi: 10.1073 / pnas.1005264107
PubMed Abstract | CrossRef Full Text/Google Scholar
Wang, W., Wang, S., Liu, Q., Wu, J., și Tao, N. (2012a). Cartografierea interacțiunilor cu o singură celulă-substrat prin microscopie de rezonanță cu plasmon de suprafață. Langmuir 28, 13373-13379. doi: 10.1021/la301712h
PubMed Abstract | CrossRef Full Text/Google Scholar
Wang, W., Yang, Y., Wang, S., Nagaraj, V. J., Liu, Q., Wu, J. și colab. (2012b). Măsurarea și cartografierea fără etichete a cineticii de legare a proteinelor membranare în celule vii unice. Nat. Chem. 4, 846–853. doi: 10.1038 / nchem.1434
PubMed Abstract | CrossRef Full Text/Google Scholar
Wang, W., Yin, L., Gonzalez-Malerva, L., Wang, S., Yu, X., Eaton, S. și colab. (2014). Cinetica de legare a receptorilor de droguri in situ în celule unice: un studiu cantitativ fără etichetă a rezistenței la medicamente antitumorale. Sci. Rep. 4: 6609. doi: 10.1038/srep06609
PubMed Abstract | CrossRef Full Text/Google Scholar
Wang, Y., Zhang, S., Xu, T., Zhang, T., Mo, Y., Liu, J. și colab. (2018). Detectarea Ultra-sensibilă și ultra-rapidă a răspunsurilor întregi ale celulelor canceroase vii neetichetate la paclitaxel cu un biosenzor pe bază de grafen. Sens. Actionat. B Chem. 263, 417–425. doi: 10.1016 / j. snb.2018.02.095
CrossRef Full Text/Google Scholar
Yanase, Y., Hiragun, T., Ishii, K., Kawaguchi, T., Yanase, T., Kawai, M. și colab. (2014). Rezonanța plasmonului de suprafață pentru diagnosticul clinic bazat pe celule. Senzori 14, 4948-4959. doi: 10.3390/s140304948
PubMed Abstract | CrossRef Full Text/Google Scholar
Yanase, Y., Hiragun, T., Kaneko, S., Gould, H. J., Greaves, M. W. și ascunde, M. (2010). Detectarea modificărilor indicelui de refracție în celulele vii individuale prin imagistica prin rezonanță a plasmonului de suprafață. Biosens. Bioelectron. 26, 674–681. doi: 10.1016/j.bios.2010.06.065
PubMed Abstract | CrossRef Full Text/Google Scholar
Yang, Y., Yu, H., Shan, X., Wang, W., Liu, X., Wang, S., și colab. (2015). Urmărirea fără etichete a transportului unic de organelle în celule cu precizie nanometrică folosind o tehnică de imagistică plasmonică. Mic 11, 2878-2884. doi: 10.1002 / smll.201403016
PubMed Abstract | CrossRef Full Text/Google Scholar
Zhang, F., Wang, S., Yin, L., Yang, Y., Guan, Y., Wang, W. și colab. (2015). Cuantificarea nivelului de expresie a receptorului factorului de creștere epidermal și a cineticii de legare pe suprafețele celulare prin imagistica prin rezonanță plasmonică de suprafață. Anal. Chem. 87, 9960–9965. doi: 10.1021 / acs.analchem.5b02572
PubMed rezumat / CrossRef textul integral / Google Scholar