16.2:ウイルスのライフサイクル
溶原性経路
いくつかの細菌ウイルスのためのより良いオプ この選択肢を有するバクテリオファージは、溶解経路と同様に、温帯ファージとして知られている。 この経路では、ウイルスは宿主ゲノムに統合することによって休眠状態に入り、環境条件が変化し、より多くの宿主細胞が感染する可能性を反映するまで転写的に静止したままである(図\(\PageIndex{4}\))。2019-01-09の8.52.35AMでスクリーンショットを撮影しました。図\(\PageIndex{4}\)を参照してください。 溶原性経路
ラムダ(λ)は、温帯バクテリオファージの一例です。 感染およびゲノム注入の初期段階は溶解サイクルと同じであるが、溶解生成を促進する条件下では、ウイルスゲノムはステップ3で宿主ゲノムに統合される。 Eに統合されている。 大腸菌では、これはatt β部位として知られる15塩基対配列での相互組換えによって起こり、Int遺伝子産物によって促進される。 環境条件が細菌の再生を助長しない限り(したがって、可能な宿主細胞の数が限られている)、ウイルスゲノムはほとんど隠されて不活性のままである。 唯一の重要な例外は、次のステップを防ぎ、ウイルスを休眠状態に保つa βリプレッサーをコードする遺伝子です。その次のステップは、宿主染色体からのλファージDNAの切除、およびその後のウイルスDNAの複製および転写である(図\(\PageIndex{4}\)、ステップ4)。 その後、前のように、最終的なステップは、ビリオンの組み立てと蓄積、および細胞構造の最終的な分解およびウイルス粒子の放出である。
カプシドによって与えられるタイトなスペース制約に包装するためには、ウイルスのゲノムは非常に経済的でなければなりません。 例えば、HIVゲノム(図\(\PageIndex{6}\))には、重複するいくつかの遺伝子があります。2019-01-09午前8時51分42秒にスクリーンショットを撮影しました。図\(\PageIndex{6}\)を参照してください。 HIVゲノム
または、curtoviruses、ssDNA植物ウイルス(例えばbeet curly top virus)の場合、ゲノムは過剰再ラッピング遺伝子を有するだけでなく、ssDNAがdsDNAに変換された後、DNAの両鎖に遺伝子産物をコードする双方向でもある(図\(\PageIndex{7}\))。2019-01-09の8.51.11AMでスクリーンショットを撮影しました。図\(\PageIndex{7}\)を参照してください。 Curtovirusゲノム
経済の必要性を考えると、ウイルスにはどのような遺伝子がありますか? 最も研究されているウイルスゲノムの一つであるバクテリオファージγには、五つの転写制御タンパク質(ファージがリゾゲンか溶解モードかに依存する)、転写活性化因子の分解を制御する結合タンパク質、17のカプシドタンパク質、宿主ゲノム内のファージゲノムの切除および挿入を制御する切除酵素、宿主ゲノムにファージゲノムを挿入するインテグレーションタンパク質、および宿主細胞の溶解に関与する3つの遺伝子が含まれている。
上記のHIVゲノムは、48kbと比較して約9キロベースで、γよりもはるかに小さいですが、再び、テーマは、可能な限り細胞タンパク質を使用し、必要に応じてウ したがって、gagはカプシド蛋白質をコードし、polは逆転写酵素、インテグラーゼ、HIVプロテアーゼ(gagおよびpol遺伝子産物を機能性蛋白質に切断する)をコードし、vifは共通の宿主細胞抗ウイルス酵素に作用し、vprは核輸入を調節し、tatはHIV遺伝子の転写を強く増加させ、revは核からウイルスRNAを輸出し、vpuは宿主からの粒子の出芽に必要であり、envはウイルスエンベロープ糖タンパク質をコードし、nefは感染細胞の生存を促進する。 LTR領域は、これらの遺伝子の高発現を駆動するための非常に強力なプロモーターである。それは溶解生成と呼ばれていませんが、いくつかの動物ウイルスは同様に行動することができます。
それは溶解生成と呼ばれていませんが、 最も顕著な例は、一般的にレトロウイルスとして知られているボルチモアクラスVIウイルスであり、そのうちの一つはHIVである。 真核生物宿主細胞を通るレトロウイルスの経路を以下に示す(g.5)。2019-01-09の8.51.19AMでスクリーンショットを撮影しました。図\(\PageIndex{5}\)を参照してください。 真核生物宿主細胞におけるレトロウイルスの感染および再生経路。
HIVにはエンベロープがあり、宿主細胞によって認識される膜貫通タンパク質がちりばめられ、ウイルスを細胞表面に結合し、受容体を介したエンドサイトーシスを開始する(1)。 エンドサイトーシスの後、ビリオンの膜エンベロープと小胞膜が融合してカプシドとその内容物を放出する(2)。 カプシドが細胞質で解離した後、ウイルスRNAの二本鎖が特別なポリメラーゼと共に放出される:RNAテンプレートを読み取り、DNAを合成する逆転写酵素。 逆転写酵素はまた、それが最終的にウイルスゲノム(の二本鎖DNAバージョンを生成するように、相補的なDNA鎖を合成するために、その新しいDNAを使用し このウイルスdsDNAは、それが別のウイルスタンパク質、インテグラーゼを使用して宿主ゲノムに統合核に輸送されます
(4)。 統合されたウイルスDNAはプロウイルスと呼ばれます。 プロウイルスは休眠状態になる可能性がありますが、活性化されると転写され、得られたウイルスRNAが核から輸送されます(5)。 ウイルスRNAの中には、逆転写酵素やインテグラーゼ、カプシドタンパク質などの酵素をコードしているものもありますが(6)、膜に結合した糖タンパク質をコードしているものもあり、これらはERに移行し(7)、最終的にゴルジを介して処理され、原形質膜に組み込まれています(9)。 ビリオンが組み立てられると(8)、ウイルス膜貫通タンパク質に結合し、ウイルスエンベロープを備えたビリオンである外細胞の”小胞”(10)を核形成する。HIVゲノムの最近の構造検査は、HIV RNA自体の構造が宿主細胞内での増殖に重要な役割を果たす可能性があることを示唆している。
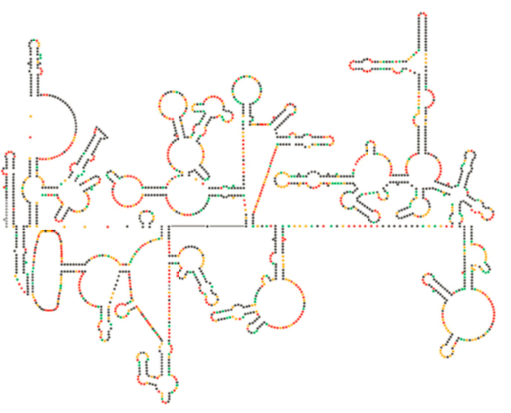
HIV rna自体の構造は、宿主細胞内での増殖に重要な役割を果たす可能性があることを示唆している。 Watts et al,Nature460:711-716,2009からの図\(\PageIndex{8}\)は、ゲノムの予測された二次構造を示している。 著者たちは、RNA構造が実際にリボソーム伸長と相互作用してウイルスタンパク質の折り畳みを制御している可能性があることを示唆している。 彼らはまた、塩基配列だけでなく、任意のRNAウイルスの二次構造および三次構造にコードされた重要な遺伝情報を含むために、この引数の拡張を仮定し
この統合的な章の残りの部分に関してウイルスを考慮する際には、心に留めておくべき二つのオーバーライドのアイデアがあります。 第一に、ウイルスの生存は数字に基づいています:それは可能な限り新しい宿主細胞のための広いネットをキャストするために、その成分の膨大な数 これを行うために、ウイルスプロモーターは通常、宿主細胞プロモーターよりもはるかに強力であり、同時により多くのウイルス遺伝子発現を駆動し、宿主遺伝子発現を防止する(細胞資源をウイルス産生に専念させることによって)。 第二に、速い生成時間のために、ウイルスの突然変異および進化の速度は、通常の真核生物のゲノムよりもはるかに速い。 さらに、ウイルスが独自のポリメラーゼ(逆転写酵素やRNA replicaseなど)を使用すると、ウイルスポリメラーゼによるエラーチェックがないため、変異率はさらに上昇する。